DNT molekulu yerləşir. DNT və genlər. DNT-yə yeni baxış
İrsiyyətin biokimyəvi əsasları.
Nuklein turşularının genetik rolu.
Nuklein turşuları ibtidaidən mürəkkəbə qədər bütün hüceyrələrdə olan bioloji polimerlərdir. Onlar ilk dəfə 1868-ci ildə İohann Fridrix Mişer tərəfindən nüvə materialı ilə zəngin hüceyrələrdə (leykositlər, qızılbalıq sperması) aşkar edilmişdir. "Nuklein turşuları" termini 1889-cu ildə təklif edilmişdir.
Nuklein turşularının iki növü var: DNT, RNT (ATP - mononükleotid). DNT və RNT molekullardır - matrislər. DNT somatik hüceyrələrdə təxminən 6 * 10 -12 g ehtiva edir: nüvədə, mitoxondriyada. RNT ribosomların bir hissəsidir və nüvədə və sitoplazmada olur.
Nuklein turşularının irsi məlumatların ötürülməsində aparıcı rolunun öyrənilməsi və sübutu virus hissəcikləri üzərində aparılmışdır. Tütün mozaika virusunun tütün və bağayarpağı üçün təhlükəli olduğu məlumdur. Viral hissəcik 95% protein və 5% nuklein turşusundan ibarətdir. Viral hissəciklərdəki protein kapsidi dəyişdirildi, lakin bir müddət sonra hər iki ştammdakı protein əvvəlki formasına çevrildi.
E. coli-ni yoluxduran bakteriofaqlarda fag qabığının zülalları radioaktiv S ilə, fag DNT-si isə radioaktiv P ilə işarələnmişdir. Fagla yoluxmuş bakteriya hüceyrəsində tərkibində yalnız radioaktiv P olan faq hissəcikləri əmələ gəlmişdir.
DNT və RNT molekullarının quruluşu və funksiyaları.
Nuklein turşuları qeyri-müntəzəm strukturlu biopolimerlərdir, monomerləri nukleotidlərdir. Nukleotid üç maddənin qalıqlarından ibarətdir: fosfor turşusu, karbohidrat - pentoza, azotlu əsas. DNT nukleotidləri karbohidrat dezoksiriboza, RNT isə riboza ehtiva edir. DNT-ni təşkil edən purin və pirimidin azotlu əsasların qalıqları adenin, quanin, sitozin, timindir. RNT molekullarında adenin, guanin, sitozin və urasil var.
Nukleotidlər bir nukleotidin fosfor turşusu qalığı və digərinin karbohidratı vasitəsilə bir-birinə “oksigen körpüsü” adlanan güclü kovalent ester bağı ilə bağlanır. Bağ bir nukleotidin karbohidratının 5-ci karbon atomundan keçərək başqa bir nukleotidin karbohidratının 3-cü karbon atomuna keçir. Nukleotid ardıcıllığı nuklein turşularının ilkin quruluşunu təmsil edir. RNT tək polinükleotid zənciridir. DNT-nin quruluşu spiral şəklində bükülmüş ikiqat polinükleotid zəncirindən ibarətdir.
DNT-nin ikincil quruluşu birinciyə nisbətən tamamlayıcılıq prinsipinə uyğun düzülmüş ikinci bir DNT zəncirinin meydana çıxması ilə əmələ gəlir. İkinci zəncir birincinin əksinədir (antiparalel). Azotlu əsaslar molekulun müstəvisinə perpendikulyar bir müstəvidə yerləşir - bu, spiral pilləkənə bənzəyir. Bu pilləkənin məhəccərləri fosfor turşusu qalıqları və karbohidratlar, pilləkənlər isə azotlu əsaslardır.
Əks istiqamətli zəncirlərdə hər bir nukleotidi təşkil edən azotlu əsaslar bir-biri ilə tamamlayıcı hidrogen bağları yaratmağa qadirdir (hər bir azotlu əsasın strukturunda mövcud olan funksional qruplara görə). Adenil nukleotidi timin, quanil sitozinə və əksinə tamamlayıcıdır. Bu bağlar özlüyündə kövrəkdir, lakin bu cür bağlarla bütün uzunluğu boyunca dəfələrlə “tikilmiş” bir DNT molekulu çox güclü bir əlaqəni təmsil edir.
Tamamlayıcılıq- bu, azotlu əsasların bir-birinə məkan-struktur və kimyəvi uyğunluğudur; onlar bir-birinə "qıfılın açarı kimi" uyğun gəlir.
Bir DNT molekulunda 10 8 və ya daha çox nukleotid ola bilər.
DNT molekulunun ikiqat antiparalel sarmal kimi quruluşu 1953-cü ildə amerikalı bioloq Ceyms Uotson və ingilis fiziki Frensis Krik tərəfindən təklif edilmişdir.
Planetdəki hər hansı canlı orqanizmin DNT molekulu tərkibində olan azotlu əsaslara görə bir-birindən fərqlənən yalnız dörd növ nukleotiddən ibarətdir: adenil, quanil, timin və sitozin. Bunda çox yönlülük DNT. Onların ardıcıllığı fərqlidir və sayı sonsuzdur.
Canlı orqanizmlərin hər bir növü və hər bir orqanizm üçün ayrıca, bu ardıcıllıq fərdi və ciddi şəkildə spesifik .
Özəllik DNT-nin quruluşu ondan ibarətdir ki, molekulun kimyəvi cəhətdən aktiv bölmələri - azotlu əsaslar, spiralın mərkəzinə batırılır və bir-biri ilə tamamlayıcı bağlar əmələ gətirir, dezoksiriboza və fosfor turşusu qalıqları isə periferiyada yerləşir və azotlu əsaslara girişi əhatə edir. - kimyəvi cəhətdən təsirsizdirlər. Bu quruluş uzun müddət kimyəvi sabitliyi saxlaya bilir. İrsi məlumatları saxlamaq üçün başqa nə lazımdır? Məhz DNT strukturunun bu xüsusiyyətləri onun genetik məlumatı kodlaşdırmaq və çoxaltmaq qabiliyyətini müəyyən edir.
DNT-nin güclü strukturunu məhv etmək olduqca çətindir. Buna baxmayaraq, bu, hüceyrədə müntəzəm olaraq baş verir - RNT sintezi və hüceyrə bölünməsindən əvvəl DNT molekulunun özünün ikiqat artması zamanı.
Duplikasiya, DNT replikasiyası xüsusi fermentin - DNT polimerazın qoşa spiralın açılması və onu ayrı-ayrı zəncirlərə ayırması - reduplikasiya çəngəlinin əmələ gəlməsi ilə başlayır. Ferment fermuarda kilid kimi fəaliyyət göstərir. Hər bir təkzəncirli zəncirdə - reduplikasiya çəngəlinin yapışqan uclarında - tamamlayıcılıq prinsipinə uyğun olaraq karioplazmada sərbəst nukleotidlərdən yeni zəncir sintez olunur. İki yeni DNT molekulunda bir zəncir orijinal ana zəncir, ikincisi isə yeni qız zəncirində qalır. Nəticədə, bir DNT molekulu yerinə, orijinalı ilə eyni dəqiq nukleotid tərkibinə malik iki molekul meydana çıxır.
Canlı sistemlərdə biz cansız təbiətdə bilinməyən yeni reaksiya növü ilə qarşılaşırıq. Onlar çağırılır matris sintez reaksiyaları . Matris sintezi matris üzərinə tökmə kimidir: yeni molekullar mövcud molekulların strukturunda göstərilən plana uyğun olaraq tam olaraq sintez edilir. Bu reaksiyalar sintez olunan polimerlərdə monomer vahidlərinin dəqiq ardıcıllığını təmin edir. Monomerlər reaksiyanın baş verdiyi matris rolunu oynayan molekullarda müəyyən bir yerə çatır. Əgər belə reaksiyalar molekulların təsadüfi toqquşması nəticəsində baş versəydi, sonsuz yavaş gedərdi. Şablon prinsipi əsasında mürəkkəb molekulların sintezi fermentlərin köməyi ilə tez və dəqiq həyata keçirilir. Şablon sintezi nuklein turşularının və zülalların sintezində ən mühüm reaksiyaların əsasını təşkil edir. Hüceyrədəki matrisin rolunu nuklein turşularının DNT və ya RNT molekulları oynayır. Polimerin sintez olunduğu monomerik molekullar - nukleotidlər və ya amin turşuları - tamamlayıcılıq prinsipinə uyğun olaraq, ciddi şəkildə müəyyən edilmiş qaydada matrisdə yerləşdirilir və sabitlənir. Sonra monomer vahidləri bir polimer zəncirinə birləşdirilir və hazır polimer matrisi tərk edir. Bundan sonra matris yeni, tam olaraq eyni polimer molekulunun yığılmasına hazırdır.
Matris tipli reaksiyalar canlı hüceyrənin spesifik xüsusiyyətidir. Onlar bütün canlıların əsas xüsusiyyətinin - öz növünü çoxaltmaq qabiliyyətinin əsasını təşkil edir.
Nuklein turşularının funksiyaları– irsi məlumatların saxlanması və ötürülməsi. DNT molekulları zülalın ilkin quruluşu haqqında məlumatları kodlayır. mRNT molekullarının sintezi DNT matrisində baş verir. Bu proses "transkripsiya" adlanır. I-RNT “tərcümə” prosesində məlumatı zülal molekulunda amin turşularının ardıcıllığı şəklində həyata keçirir.
Hər bir hüceyrənin DNT-si təkcə hüceyrənin formasını təyin edən struktur zülallar haqqında deyil, həm də bütün ferment zülalları, hormon zülalları və digər zülallar, eləcə də bütün növ RNT-lərin strukturu haqqında məlumat daşıyır.
Ola bilsin ki, nuklein turşuları bioloji yaddaşın müxtəlif növlərini - immunoloji, nevroloji və s. təmin edir, həmçinin biosintetik proseslərin tənzimlənməsində mühüm rol oynayır.
Əlaqədar məlumat.
DNT-nin orqanizmlərdə irsi məlumat daşıyıcısı kimi rolunun ilk sübutu nuklein turşularının öyrənilməsinə böyük diqqət yetirdi. 1869-cu ildə F.Mişer hüceyrə nüvələrindən nuklein adlandırdığı xüsusi maddə ayırdı. 20 ildən sonra bu ad terminlə əvəz olundu nuklein turşusu. 1924-cü ildə R.Felgen nuklein turşularının spesifik boyanma yolu ilə sitoloji tanınması metodunu işləyib hazırladı və göstərdi ki, DNT hüceyrələrin nüvələrində, RNT isə sitoplazmada lokallaşdırılır. 1936-cı ildə A.N. Belozersky və I.I. Dubrovskaya bitki hüceyrələrinin nüvələrindən təmiz formada DNT-ni təcrid etdi. 1930-cu illərin əvvəllərində. Nuklein turşusu şəkərlərinin quruluşunun əsas kimyəvi prinsipləri aydınlaşdırıldı və 1953-cü ildə DNT-nin struktur modeli yaradıldı.
Nuklein turşularının əsas struktur vahidi nukleotid, kovalent bağlarla birləşdirilmiş üç kimyəvi cəhətdən fərqli hissədən ibarətdir (şək. 5.2).
düyü. 5.2. Struktur formullar: A- nukleotidlər; b- DNT; V - RNT (həmçinin bax, səh. 110)

düyü. 5.2. Bitiş. Struktur formullar: A- nukleotidlər; 6 - DNT; V- RNT
Birinci hissə beş karbon atomu olan şəkərdir: deoksiriboza DNT-də və riboza RNT-də.
Nukleotidin ikinci hissəsi, purin və ya pirimidin azotlu əsas, şəkərin birinci karbon atomu ilə kovalent şəkildə bağlıdır və adlanan bir quruluş meydana gətirir. nukleozid. DNT-də purin əsasları var - adenin(A) və guanin(D) - və pirimidin əsasları - timin(T) və sitozin(C). Müvafiq nukleozidlər deoksiadenozin, deoksiquanozin, deoksitimidin və deoksisitidin adlanır. RNT, pirimidin əsası olan DNT ilə eyni purin əsaslarını ehtiva edir sitozin, və timin əvəzinə ehtiva edir urasil(U); müvafiq nukleozidlər adenozin, quanozin, uridin və sitidin adlanır.
Nukleotidin üçüncü hissəsi bir şəkərin 5 karbon atomu ilə digərinin 3 karbon atomu arasında fosfodiester bağları vasitəsilə qonşu nukleozidləri polimer zəncirinə birləşdirən fosfat qrupudur (şək. 5.2, b, V). Nukleotidlərşəkərin 3" və ya 5 karbon atomlarına ester bağları ilə bağlanmış bir və ya bir neçə fosfat qrupu olan nukleozidlər adlanır. Nukleotidlərin sintezi nuklein turşularının sintezindən əvvəl olur; müvafiq olaraq, nukleotidlər nuklein turşularının kimyəvi və ya fermentativ hidrolizinin məhsullarıdır.
Nuklein turşuları 5 və 3'-fosfodiester bağları ilə bağlanmış mononükleotidlərdən ibarət çox uzun polimer zəncirləridir. Bütöv DNT molekulunda orqanizmin növündən asılı olaraq bir neçə mindən çox milyonlarla nukleotid, bütöv bir RNT molekulu - 100 ilə 100 min və ya daha çox nukleotid var.
E.Çarqaffın müxtəlif növ formalı DNT-nin nukleotid tərkibinin təhlilinin nəticələri göstərdi ki, müxtəlif azotlu əsasların - adenin, quanin, timin, sitozinin molekulyar nisbəti geniş şəkildə dəyişir. Nəticədə, DNT-nin 40-cı illərdə güman edildiyi kimi, eyni tetranukleotidlərdən ibarət monoton polimer olmadığı sübut edildi. XX əsr və o, nukleotid əsaslarının müəyyən bir ardıcıllığı şəklində irsi məlumatın qorunması və ötürülməsi üçün lazım olan mürəkkəbliyə tam malikdir.
E.Çarqaffın tədqiqatı bütün DNT molekullarına xas olan bir xüsusiyyəti də aşkar etdi: adeninin molyar tərkibi timin tərkibinə, quaninin molyar tərkibi isə sitozinin tərkibinə bərabərdir. Bu bərabərliklər Çarqaffın ekvivalentlik qaydası adlanır: [A] = [T], [G] = [C]; purinlərin sayı pirimidinlərin sayına bərabərdir. Növlərdən asılı olaraq yalnız ([A] + [T])/([G] + [C]) nisbəti dəyişir (Cədvəl 5.1).
|
Bazaların tərkibi |
Münasibət |
Asimmetriya |
|||||
|
səbəblər |
|||||||
|
(A + T)/(G + C) |
|||||||
|
Heyvanlar |
|||||||
|
Tısbağa |
|||||||
|
dəniz xərçəngi |
|||||||
|
Dənizkirpisi |
|||||||
|
Bitkilər, göbələklər |
|||||||
|
buğda toxumu |
|||||||
|
Göbələk Aspergillus niger |
|||||||
|
Bakteriya |
|||||||
|
Escherichia coli |
|||||||
|
Staphylococcus aureus |
|||||||
|
Clostridium perfringens |
|||||||
|
Brucela abort |
|||||||
|
Sarcina lutea |
|||||||
|
Bakteriofaqlar |
|||||||
|
FH 174 (viral forma) |
|||||||
|
FH 174 (replikativ forma) |
|||||||
Əsasların nisbəti deyilir nukleotid əmsalı(növ) spesifiklik.Çarqafın kəşfi DNT-nin mühüm struktur xüsusiyyətini formalaşdırdı və bu, daha sonra J. Watson və F. Crick (1953) tərəfindən DNT-nin struktur modelində öz əksini tapdı, o, əslində Chargaff qaydalarının mümkün birləşmələrin mümkün sayına heç bir məhdudiyyət qoymadığını göstərdi. molekulların DNT əmələ gətirə bilən müxtəlif əsas ardıcıllığı.
Nukleotid spesifikliyi konsepsiyası biologiyanın yeni bir sahəsinin əsasını təşkil etdi - gen sistematikası, orqanizmlərin təbii sistemini qurmaq üçün nuklein turşularının tərkibini və quruluşunu müqayisə etməklə fəaliyyət göstərir.
Watson-Crick modelinə görə, bir DNT molekulu tamamlayıcı prinsipə uyğun olaraq azotlu əsaslar arasında eninə hidrogen bağlarından istifadə edərək bir-birinə bağlanan iki polinükleotid zəncirindən (zəncir, zəncir) ibarətdir (bir zəncirin adenini iki hidrogen bağı ilə bağlanır). əks zəncirin timin və müxtəlif zəncirlərin guanin və sitozin bir-birinə üç hidrogen bağı ilə bağlıdır). Bu halda, bir molekulun iki polinükleotid zənciri antiparaleldir, yəni bir zəncirin 3" ucunun qarşısında digər zəncirin 5" ucu və əksinədir (şək. 5.3). Bununla belə, müasir məlumatları nəzərə almaq lazımdır ki, bəzi virusların genetik materialı tək zəncirli (tək zəncirli) DNT molekulları ilə təmsil olunur. C.Uotson və F.Krik DNT-nin rentgen şüalarının difraksiya analizinə əsaslanaraq, həmçinin belə nəticəyə gəldilər ki, onun qoşa zəncirli molekulu soldan sağa doğru bükülmüş spiral şəklində ikinci dərəcəli quruluşa malikdir və bu, sonradan 5 adlanır. -forma (şək. 5.4). Bu günə qədər sübut edilmişdir ki, ən çox yayılmış 5 formaya əlavə olaraq, fərqli konfiqurasiyaya malik olan DNT bölmələrini - sağ əlli (formalar) aşkar etmək mümkündür. A, C) və sağdan sola bükülmüş (sola bükülmüş və ya Z şəklində) (Şəkil 5.4). İkinci dərəcəli DNT strukturunun bu formaları arasında müəyyən fərqlər vardır (Cədvəl 5.2). Məsələn, nanometrlərlə (nm) ifadə edilən ikiqat sarmalda iki bitişik cüt azotlu əsaslar arasındakı məsafə 5 forması və Z forması (müvafiq olaraq 0,34 və 0,38 nm) üçün fərqli dəyərlərlə xarakterizə olunur. . Şəkildə. Şəkil 5.5-də DNT-nin “sol əlli” və “sağ əlli” formalarının müasir üçölçülü modelləri göstərilir.

düyü. 5.3. ikiqat zəncirli DNT molekulunun fraqmentinin ilkin strukturunun sxematik təsviri: A - adenin; G - guanin; T - timin; C - sitozin

düyü. 5.4.

Cədvəl 5.2
DNT cüt spirallarının müxtəlif formalarının xüsusiyyətləri
RNT molekulları struktur və funksional xüsusiyyətlərindən asılı olaraq bir neçə növə bölünür: xəbərçi RNT (mRNT və ya mRNT), ribosomal RNT (rRNT), transfer RNT (tRNT), kiçik nüvə RNT (snRNA) və s. DNT, RNT molekulları həmişə tək zəncirlidir (tək zəncirlidir). Bununla belə, onlar tamamlayıcı azotlu əsasların (A-U və G-C) qarşılıqlı təsiri əsasında belə zəncirin ayrı-ayrı hissələrinin tamamlayıcı əlaqəsi hesabına daha mürəkkəb (ikinci dərəcəli) konfiqurasiyalar yarada bilirlər. Nümunə olaraq, fenilalanin transfer RNT molekulu üçün yonca yarpağı konfiqurasiyasını nəzərdən keçirək (Şəkil 5.6).

düyü. 5.6.
1953-cü ildə D.Watson və F.Crick DNT-nin quruluşunun modelini təklif etdilər və bu model aşağıdakı postulatlara əsaslanır:
- 1. DNT 3" və 5" fosfodiester bağları ilə bağlanmış nukleotidlərdən ibarət polimerdir.
- 2. DNT nukleotidlərinin tərkibi Chargaff qaydalarına tabedir.
- 3. DNT molekulu spiral pilləkəni xatırladan ikiqat spiral quruluşa malikdir, bunu ilk dəfə M. Wilkins və R. Franklin tərəfindən əldə edilmiş DNT zəncirlərinin rentgen difraksiya nümunələri sübut edir.
- 4. Doğma (təbii) DNT-nin turşu-qələvi titrlənməsi ilə göstərildiyi kimi polimerin strukturu hidrogen bağları ilə sabitləşir. Doğma DNT-nin titrlənməsi və qızdırılması onun fiziki xassələrində, xüsusən də özlülüyündə nəzərəçarpacaq dəyişikliyə səbəb olur, onu denatürasiya edilmiş formaya çevirir və kovalent bağlar məhv edilmir.
Nuklein turşuları 3", 5" fosfodiester bağlarından istifadə edərək bir-birinə polimer zəncirində bağlanan və hüceyrələrdə müəyyən şəkildə qablaşdırılan mononükleotidlərdən ibarət yüksək molekullu maddələrdir.
Nuklein turşuları iki növ biopolimerdir: ribonuklein turşusu (RNT) və deoksiribonuklein turşusu (DNT). Hər bir biopolimer karbohidrat qalığı (riboza, dezoksiriboza) və azotlu əsaslardan biri (urasil, timin) ilə fərqlənən nukleotidlərdən ibarətdir. Bu fərqlərə görə, nuklein turşuları öz adlarını aldılar.
Dezoksiribonuklein turşusunun quruluşu
Nuklein turşuları ilkin, ikincili və üçüncülü quruluşa malikdir.
DNT-nin ilkin quruluşu
DNT-nin əsas strukturu mononükleotidlərin 3 "5" fosfodiester bağları ilə bağlandığı xətti polinükleotid zənciridir. Hüceyrədə nuklein turşusu zəncirinin yığılması üçün başlanğıc material β və γ fosfor turşusu qalıqlarının çıxarılması nəticəsində başqa bir nukleozidin 3" karbon atomunu birləşdirməyə qadir olan 5"-trifosfat nukleoziddir. . Beləliklə, bir dezoksiribozun 3" karbon atomu bir fosfor turşusu qalığı vasitəsilə digər dezoksiribozun 5" karbon atomu ilə kovalent şəkildə bağlanır və nuklein turşusunun xətti polinükleotid zəncirini əmələ gətirir. Beləliklə, adı: 3", 5" fosfodiester bağları. Azot əsasları bir zəncirin birləşən nukleotidlərində iştirak etmir (şək. 1.).
Bir nukleotidin fosfor turşusu molekulu qalığı ilə digərinin karbohidratı arasındakı belə bir əlaqə, azotlu əsasların bir-birinin ardınca yan tərəfə bağlandığı polinükleotid molekulunun pentoza-fosfat skeletinin meydana gəlməsinə səbəb olur. Onların nuklein turşusu molekullarının zəncirlərində düzülmə ardıcıllığı müxtəlif orqanizmlərin hüceyrələri üçün ciddi şəkildə spesifikdir, yəni. spesifik xarakterə malikdir (Çarqaff qaydası).
Uzunluğu zəncirə daxil olan nukleotidlərin sayından asılı olan xətti DNT zəncirinin iki ucu var: biri 3" ucu adlanır və sərbəst hidroksil ehtiva edir, digəri isə 5" ucu adlanır və fosfor ehtiva edir. turşu qalığı. Dövrə qütbdür və istiqaməti 5"->3" və 3"->5" ola bilər. İstisna dairəvi DNT-dir.
DNT-nin genetik "mətni" kod "sözlərindən" - kodon adlanan üçlü nukleotidlərdən ibarətdir. Bütün növ RNT-nin ilkin strukturu haqqında məlumatı ehtiva edən DNT bölmələrinə struktur genlər deyilir.
Polinukleotid DNT zəncirləri nəhəng ölçülərə çatır, buna görə də hüceyrədə müəyyən bir şəkildə qablaşdırılır.
Chargaff (1949) DNT-nin tərkibini öyrənərkən fərdi DNT əsaslarının məzmunu ilə bağlı mühüm qanunauyğunluqlar yaratmışdır. Onlar DNT-nin ikincil quruluşunu aşkar etməyə kömək etdilər. Bu nümunələrə Chargaff qaydaları deyilir. Chargaff qaydaları
Bu qaydalar göstərir ki, DNT-nin qurulması zamanı purin və pirimidin əsaslarının deyil, xüsusi olaraq timin ilə adenin və sitozin ilə guaninin kifayət qədər ciddi uyğunluğu (qoşulması) müşahidə edilməlidir. Bu qaydalara əsaslanaraq 1953-cü ildə Watson və Crick DNT-nin ikinci dərəcəli strukturunun qoşa spiral adlanan modelini təklif etdilər (şək.). |

DNT-nin ikincil quruluşu
DNT-nin ikincil strukturu ikiqat sarmaldır, onun modeli 1953-cü ildə D.Vatson və F.Krik tərəfindən təklif edilmişdir.
DNT modelinin yaradılması üçün ilkin şərtlər
İlkin təhlillər nəticəsində hər hansı mənşəli DNT-də bərabər molyar miqdarda dörd nukleotidin hamısının olduğu güman edilirdi. Lakin 1940-cı illərdə E.Çarqaff və onun həmkarları müxtəlif orqanizmlərdən təcrid olunmuş DNT-nin təhlili nəticəsində onların tərkibində müxtəlif kəmiyyət nisbətlərində azotlu əsaslar olduğunu açıq şəkildə göstərmişlər. Chargaff müəyyən etdi ki, bu nisbətlər eyni növ orqanizmin bütün hüceyrələrinin DNT-si üçün eyni olsa da, müxtəlif növlərdən olan DNT müəyyən nukleotidlərin tərkibində əhəmiyyətli dərəcədə fərqlənə bilər. Bu, azotlu əsasların nisbətindəki fərqlərin bir növ bioloji kodla əlaqəli ola biləcəyini göstərirdi. Fərqli DNT nümunələrində fərdi purin və pirimidin əsaslarının nisbətinin fərqli olduğu ortaya çıxsa da, test nəticələrini müqayisə edərkən müəyyən bir nümunə ortaya çıxdı: bütün nümunələrdə purinlərin ümumi sayı pirimidinlərin ümumi sayına bərabər idi (A + G = T + C), adeninin miqdarı timin miqdarına (A = T), guaninin miqdarı isə sitozinin miqdarına (G = C) bərabər idi. Məməli hüceyrələrindən təcrid olunmuş DNT ümumiyyətlə adenin və timinlə daha zəngin, guanin və sitozində isə nisbətən yoxsul idi, halbuki bakteriyalardan alınan DNT guanin və sitozinlə daha zəngin, adenin və timinlə isə nisbətən yoxsul idi. Bu məlumatlar sonradan DNT strukturunun Watson-Crick modelinin qurulduğu faktiki materialın mühüm hissəsini təşkil edirdi.
DNT-nin mümkün strukturunun digər mühüm dolayı göstəricisi L. Paulinqin zülal molekullarının quruluşu haqqında məlumatları ilə təmin edilmişdir. Pauling göstərdi ki, zülal molekulunda amin turşusu zəncirinin bir neçə müxtəlif sabit konfiqurasiyası mümkündür. Ümumi peptid zəncirinin konfiqurasiyası, α-sarmal, müntəzəm spiral quruluşdur. Bu quruluşla zəncirin bitişik döngələrində yerləşən amin turşuları arasında hidrogen bağlarının əmələ gəlməsi mümkündür. Pauling 1950-ci ildə polipeptid zəncirinin α-spiral konfiqurasiyasını təsvir etdi və DNT molekullarının, ehtimal ki, hidrogen bağları ilə yerində saxlanılan spiral quruluşa malik olduğunu irəli sürdü.
Bununla belə, DNT molekulunun quruluşu haqqında ən qiymətli məlumat rentgen şüalarının difraksiya analizinin nəticələri ilə təmin edilmişdir. DNT kristalından keçən rentgen şüaları difraksiyaya məruz qalır, yəni müəyyən istiqamətlərdə əyilir. Şüaların əyilmə dərəcəsi və xarakteri molekulların özlərinin quruluşundan asılıdır. X-şüalarının difraksiya nümunəsi (şəkil 3) təcrübəli gözə öyrənilən maddənin molekullarının quruluşu ilə bağlı bir sıra dolayı göstəricilər verir. DNT-nin rentgen şüalarının difraksiya nümunələrinin təhlili belə nəticəyə gəldi ki, azotlu əsaslar (düz formaya malik olan) lövhələr yığını kimi düzülüblər. X-şüalarının difraksiya nümunələri kristal DNT-nin strukturunda üç əsas dövrü aşkar etdi: 0,34, 2 və 3,4 nm.

Watson-Crick DNT modeli
Çarqaffın analitik məlumatlarına, Uilkinsin rentgen sxemlərinə və molekuldakı atomlar arasındakı dəqiq məsafələr, müəyyən bir atomun bağları arasındakı bucaqlar və atomların ölçüsü haqqında məlumat verən kimyaçıların araşdırmalarına əsaslanaraq, Watson və Crick müəyyən miqyasda DNT molekulunun ayrı-ayrı komponentlərinin fiziki modellərini qurmağa və onları bir-birinə elə "tənzimləməyə" başladı ki, nəticədə yaranan sistem müxtəlif eksperimental məlumatlara uyğun olsun. [göstərmək] .
DNT zəncirindəki qonşu nukleotidlərin bir nukleotidin 5"-karbon dezoksiriboza atomunu növbəti nukleotidin 3"-karbon dezoksiriboza atomu ilə birləşdirən fosfodiester körpüləri ilə bağlandığı daha əvvəl məlum idi. Watson və Crick 0,34 nm müddətinin DNT zəncirindəki ardıcıl nukleotidlər arasındakı məsafəyə uyğun olduğuna şübhə etmirdilər. Bundan əlavə, 2 nm dövrünün zəncirin qalınlığına uyğun olduğunu güman etmək olar. Və 3,4 nm dövrünün hansı real quruluşa uyğun olduğunu izah etmək üçün Watson və Crick, eləcə də Pauling əvvəllər zəncirin spiral şəklində bükülməsini (daha doğrusu, spiral xətt təşkil etdiyini) təklif etdi. bu sözlərin ciddi mənasında bir spiral, rulonlar kosmosda silindrik deyil, konusvari bir səth meydana gətirdikdə əldə edilir). Sonra 3,4 nm-lik bir dövr bu spiralın ardıcıl dönüşləri arasındakı məsafəyə uyğun olacaq. Belə bir spiral çox sıx və ya bir qədər uzanmış ola bilər, yəni onun dönüşləri düz və ya dik ola bilər. 3,4 nm-lik dövr ardıcıl nukleotidlər (0,34 nm) arasındakı məsafədən düz 10 dəfə çox olduğundan, spiralın hər tam dönüşündə 10 nukleotid olduğu aydındır. Bu məlumatlara əsasən, Watson və Crick, 2 nm diametrli, döngələr arasında məsafə 3,4 nm olan bir spiral şəklində bükülmüş polinükleotid zəncirinin sıxlığını hesablaya bildilər. Məlum oldu ki, belə bir zəncirin artıq məlum olan DNT-nin həqiqi sıxlığının yarısı qədər sıxlıq olacaq. Güman etməli idim ki, DNT molekulu iki zəncirdən ibarətdir - o, nukleotidlərdən ibarət ikiqat spiraldır.
Növbəti vəzifə, əlbəttə ki, ikiqat sarmal meydana gətirən iki zəncir arasındakı məkan əlaqələrini aydınlaşdırmaq idi. Fiziki modellərində zəncirlərin düzülməsi üçün bir sıra variantları sınayan Watson və Crick tapdılar ki, bütün mövcud məlumatlar iki polinükleotid sarmalının əks istiqamətə getdiyi variantla ən yaxşı şəkildə uyğun gəlir; bu zaman şəkər və fosfat qalıqlarından ibarət zəncirlər qoşa spiralın səthini əmələ gətirir, içərisində isə purinlər və pirimidinlər yerləşir. İki zəncirə aid olan bir-birinə qarşı yerləşən əsaslar hidrogen bağları ilə cüt-cüt birləşir; Zəncirləri bir yerdə saxlayan və beləliklə molekulun ümumi konfiqurasiyasını sabitləyən bu hidrogen bağlarıdır.
DNT-nin qoşa spiralını, pilləkənləri üfüqi vəziyyətdə qalması üçün spiralvari şəkildə bükülmüş ip nərdivan kimi təsəvvür etmək olar. Sonra iki uzununa iplər şəkər və fosfat qalıqlarının zəncirlərinə, çarpaz çubuqlar isə hidrogen bağları ilə bağlanmış azotlu əsasların cütlərinə uyğun olacaq.
Mümkün modellərin sonrakı tədqiqi nəticəsində Watson və Crick belə nəticəyə gəldilər ki, hər bir "çarpaz" bir purin və bir pirimidindən ibarət olmalıdır; 2 nm müddətində (qoşa spiralın diametrinə uyğundur) iki purin üçün kifayət qədər yer olmayacaq və iki pirimidin müvafiq hidrogen bağları yaratmaq üçün bir-birinə kifayət qədər yaxın ola bilməz. Təfərrüatlı modelin dərindən tədqiqi adenin və sitozinin uyğun ölçüdə bir birləşmə meydana gətirərkən, hələ də aralarında hidrogen bağları meydana gətirəcək şəkildə yerləşdirilə bilməyəcəyini göstərdi. Oxşar hesabatlar guanin - timin birləşməsini istisna etməyə məcbur etdi, adenin - timin və guanin - sitozin birləşmələri isə olduqca məqbul olduğu ortaya çıxdı. Hidrogen bağlarının təbiəti elədir ki, adenin timinlə, guanin isə sitozinlə cüt əmələ gətirir. Bu spesifik baza cütləşməsi ideyası "Chargaff qaydasını" izah etməyə imkan verdi, buna görə hər hansı bir DNT molekulunda adeninin miqdarı həmişə timin tərkibinə, guaninin miqdarı isə həmişə miqdarına bərabərdir. sitozin. Adenin və timin arasında iki, guanin və sitozin arasında üç hidrogen bağı əmələ gəlir. Bu spesifikliyə görə bir zəncirdə hər bir adeninə qarşı hidrogen bağlarının meydana gəlməsi digərində timin meydana gəlməsinə səbəb olur; eyni şəkildə hər guaninin qarşısında yalnız sitozin ola bilər. Beləliklə, zəncirlər bir-birini tamamlayır, yəni bir zəncirdəki nukleotidlərin ardıcıllığı digərində onların ardıcıllığını unikal şəkildə müəyyən edir. İki zəncir əks istiqamətdə hərəkət edir və onların son fosfat qrupları qoşa sarmalın əks uclarındadır.
Tədqiqatları nəticəsində 1953-cü ildə Watson və Crick DNT molekulunun strukturunun modelini təklif etdilər (şək. 3), bu, günümüz üçün də aktuallığını qoruyur. Modelə görə, DNT molekulu iki tamamlayıcı polinükleotid zəncirindən ibarətdir. Hər bir DNT ipi bir neçə on minlərlə nukleotiddən ibarət polinükleotiddir. Orada qonşu nukleotidlər fosfor turşusu qalığı ilə dezoksiribozun güclü kovalent bağla bağlanması hesabına müntəzəm pentoza-fosfat onurğasını əmələ gətirir. Bir polinükleotid zəncirinin azotlu əsasları digərinin azotlu əsaslarının əksinə ciddi şəkildə müəyyən edilmiş qaydada düzülür. Polinükleotid zəncirində azotlu əsasların növbələşməsi qeyri-müntəzəmdir.
DNT zəncirində azotlu əsasların düzülüşü tamamlayıcıdır (yunan dilindən "tamamlayıcı" - əlavə), yəni. Timin (T) həmişə adeninə (A), yalnız sitozin (C) guaninə (G) qarşıdır. Bu, A və T, eləcə də G və C-nin bir-birinə ciddi şəkildə uyğun olması ilə izah olunur, yəni. bir-birini tamamlayır. Bu uyğunluq purin və pirimidin cütlüyündə hidrogen bağlarının yaranmasına imkan verən əsasların kimyəvi quruluşu ilə müəyyən edilir. A və T arasında iki, G və C arasında isə üç əlaqə var. Bu bağlar kosmosda DNT molekulunun qismən sabitləşməsini təmin edir. İkiqat sarmalın sabitliyi A=T istiqrazları ilə müqayisədə daha sabit olan G≡C bağlarının sayı ilə birbaşa mütənasibdir.
Bir DNT zəncirində nukleotidlərin məlum düzülüşü, tamamlayıcılıq prinsipinə əsasən, başqa bir zəncirin nukleotidlərini yaratmağa imkan verir.
Bundan əlavə, müəyyən edilmişdir ki, sulu məhlulda aromatik quruluşa malik olan azotlu əsaslar bir-birinin üstündə yerləşərək, sanki bir sikkə yığını əmələ gətirir. Üzvi molekulların yığınlarının əmələ gəlməsi prosesinə stacking deyilir. Baxılan Watson-Crick modelinin DNT molekulunun polinükleotid zəncirləri oxşar fiziki-kimyəvi vəziyyətə malikdir, onların azotlu əsasları müstəviləri arasında van der Waals qarşılıqlı təsirləri (yığınlaşma qarşılıqlı təsirləri) yaranan sikkələr yığını şəklində yerləşdirilir.
Tamamlayıcı əsaslar (üfüqi) və van der Waals qüvvələri (şaquli) səbəbindən bir polinükleotid zəncirindəki əsasların müstəviləri arasında yığılan qarşılıqlı təsirlər DNT molekulunun kosmosda əlavə sabitləşməsini təmin edir.
Hər iki zəncirin şəkər fosfat onurğaları xaricə, əsasları isə içəriyə, bir-birinə baxır. DNT-dəki zəncirlərin istiqaməti antiparaleldir (onlardan birinin istiqaməti 5"->3", digəri - 3"->5", yəni bir zəncirin 3" ucu 5" ucunun qarşısında yerləşir. başqa.). Zəncirlər ümumi oxlu sağ əlli spirallər əmələ gətirir. Spiralın bir növbəsi 10 nukleotiddir, növbənin ölçüsü 3,4 nm, hər bir nukleotidin hündürlüyü 0,34 nm, spiralın diametri 2,0 nm-dir. Bir zəncirinin digəri ətrafında fırlanması nəticəsində DNT cüt spiralının əsas yivi (diametri təxminən 20 Å) və kiçik yiv (diametri təxminən 12 Å) əmələ gəlir. Watson-Crick qoşa spiralının bu forması sonralar B forması adlanır. Hüceyrələrdə DNT adətən ən sabit olan B şəklində mövcuddur.
DNT-nin funksiyaları
Təklif olunan model deoksiribonuklein turşusunun bir çox bioloji xüsusiyyətlərini, o cümlədən genetik məlumatın saxlanmasını və 4 nukleotidin müxtəlif ardıcıl birləşmələri ilə təmin edilən genlərin müxtəlifliyini və genetik kodun mövcudluğu faktını, özünü çoxalma qabiliyyətini izah etdi. və təkrarlama prosesi ilə təmin edilən genetik məlumatın ötürülməsi və genetik məlumatın zülallar şəklində həyata keçirilməsi, həmçinin ferment zülallarının köməyi ilə əmələ gələn hər hansı digər birləşmələr.
DNT-nin əsas funksiyaları.
- DNT genetik kodun mövcudluğu faktı ilə təmin edilən genetik məlumatın daşıyıcısıdır.
- Genetik məlumatın hüceyrə və orqanizmlərin nəsilləri arasında çoxalması və ötürülməsi. Bu funksiya replikasiya prosesi ilə təmin edilir.
- Genetik məlumatın zülallar, həmçinin ferment zülallarının köməyi ilə əmələ gələn hər hansı digər birləşmələr şəklində həyata keçirilməsi. Bu funksiya transkripsiya və tərcümə prosesləri ilə təmin edilir.

İki zəncirli DNT-nin təşkili formaları
DNT bir neçə növ cüt spiral yarada bilər (şəkil 4). Hal-hazırda altı forma artıq məlumdur (A-dan E və Z-formasına).
DNT-nin struktur formaları, Rozalind Franklinin müəyyən etdiyi kimi, nuklein turşusu molekulunun su ilə doymasından asılıdır. X-şüalarının difraksiya analizindən istifadə edərək DNT lifləri üzərində aparılan tədqiqatlarda göstərilmişdir ki, rentgen nümunəsi eksperimentin bu lifin su ilə doyma dərəcəsinin hansı dərəcədə nisbi rütubətdən kökündən asılıdır. Lif kifayət qədər su ilə doymuşdursa, bir rentgenoqrafiya alındı. Quruduqda, yüksək nəmlik lifinin rentgen nümunəsindən çox fərqli, tamamilə fərqli bir rentgen nümunəsi meydana çıxdı.
Yüksək rütubətli DNT molekuluna B forması deyilir. Fizioloji şəraitdə (aşağı duz konsentrasiyası, yüksək nəmlənmə dərəcəsi) DNT-nin dominant struktur tipi B-formadır (ikiqat zəncirli DNT-nin əsas forması - Watson-Crick modeli). Belə bir molekulun spiral hündürlüyü 3,4 nm-dir. Bükülmüş "sikkələr" - azotlu əsaslar şəklində bir növbədə 10 tamamlayıcı cüt var. Yığınlar yığınların bir-birinə zidd olan iki "sikkəsi" arasında hidrogen bağları ilə bir yerdə saxlanılır və sağ əlli spiral şəklində bükülmüş iki fosfodiester magistral lenti ilə "sarılır". Azotlu əsasların müstəviləri spiral oxuna perpendikulyardır. Qonşu tamamlayıcı cütlər bir-birinə nisbətən 36° fırlanır. Spiralın diametri 20Å, purin nukleotidi 12Å və pirimidin nukleotidi 8Å tutur.
Aşağı rütubətli DNT molekulu A-forma adlanır. A forması daha az yüksək nəmləndirmə şəraitində və Na + və ya K + ionlarının daha çox tərkibində əmələ gəlir. Bu daha geniş sağ əlli spiral konformasiyada hər növbədə 11 əsas cüt var. Azotlu əsasların müstəviləri spiral oxuna daha güclü meylə malikdir, onlar normaldan spiral oxuna 20° kənara çıxırlar. Bu, 5Å diametrli daxili boşluğun mövcudluğunu nəzərdə tutur. Qonşu nukleotidlər arasındakı məsafə 0,23 nm, döngənin uzunluğu 2,5 nm, spiralın diametri isə 2,3 nm-dir.
DNT-nin A formasının əvvəlcə daha az əhəmiyyətli olduğu düşünülürdü. Lakin sonradan məlum oldu ki, DNT-nin A forması, B forması kimi, çox böyük bioloji əhəmiyyətə malikdir. Şablon-primer kompleksindəki RNT-DNT spiralının A forması, həmçinin RNT-RNT spiral və RNT saç tıxac strukturları (ribozun 2'-hidroksil qrupu RNT molekullarının B formasını əmələ gətirməsinə mane olur). DNT-nin A forması sporlarda olur. Müəyyən edilmişdir ki, DNT-nin A forması B formasından UB şüalarına 10 dəfə daha davamlıdır.
A-forma və B-forma DNT-nin kanonik formaları adlanır.
C-E formaları həm də sağ əllidirlər, onların əmələ gəlməsini yalnız xüsusi təcrübələrdə müşahidə etmək olar və görünür, onlar in vivo mövcud deyillər. DNT-nin C forması B DNT-yə bənzər bir quruluşa malikdir. Hər növbədə əsas cütlərin sayı 9,33, spiral növbəsinin uzunluğu 3,1 nm-dir. Baza cütləri oxa perpendikulyar mövqeyə nisbətən 8 dərəcə bir açı ilə meyllidirlər. Yivlər ölçülərinə görə B-DNT-nin yivlərinə bənzəyir. Bu halda, əsas yiv bir qədər dayaz, kiçik yiv isə daha dərindir. Təbii və sintetik DNT polinükleotidləri C formasına çevrilə bilər.
| Cədvəl 1. DNT strukturlarının bəzi növlərinin xüsusiyyətləri | |||
| Spiral tip | A | B | Z |
| Spiral meydança | 0,32 nm | 3.38 nm | 4.46 nm |
| Spiral bükülmə | Sağ | Sağ | Sol |
| Döngə başına əsas cütlərin sayı | 11 | 10 | 12 |
| Baza təyyarələri arasındakı məsafə | 0,256 nm | 0,338 nm | 0,371 nm |
| Qlikozid bağ konformasiyası | anti | anti | anti-C günah-G |
| Furanoz halqasının uyğunlaşması | C3"-endo | C2"-endo | C3"-endo-G C2"-endo-C |
| Groove eni, kiçik/böyük | 1,11/0,22 nm | 0,57/1,17 nm | 0,2/0,88 nm |
| Yiv dərinliyi, kiçik/böyük | 0,26/1,30 nm | 0,82/0,85 nm | 1,38/0,37 nm |
| Spiral diametri | 2,3 nm | 2.0 nm | 1,8 nm |
DNT-nin struktur elementləri
(qeyri-kanonik DNT strukturları)
DNT-nin struktur elementlərinə bəzi xüsusi ardıcıllıqla məhdudlaşan qeyri-adi strukturlar daxildir:
|

Z formalı DNT 1979-cu ildə d(CG)3 - heksanukleotidini öyrənərkən aşkar edilmişdir. Onu MIT professoru Alexander Rich və onun həmkarları kəşf ediblər. Z-forma DNT-nin ən mühüm struktur elementlərindən birinə çevrilmişdir, çünki onun əmələ gəlməsi DNT bölgələrində purinlərin pirimidinlərlə (məsələn, 5'-GCGCGC-3') növbələşdiyi və ya təkrarlanan 5-də müşahidə edilmişdir. '-CGCGCG-3' tərkibində metilləşdirilmiş sitozin. Z-DNT-nin əmələ gəlməsi və sabitləşməsi üçün vacib şərt sin konformasiyasında onun tərkibində purin nukleotidlərinin olması, anti-konformasiyada pirimidin əsasları ilə əvəzlənməsi idi.
Təbii DNT molekulları (CG)n kimi ardıcıllıqları ehtiva etmədikcə, əsasən sağ əlli B şəklində mövcuddur. Bununla belə, əgər belə ardıcıllıqlar DNT-nin bir hissəsidirsə, onda bu bölmələr, məhlulun ion gücü və ya fosfodiester çərçivəsindəki mənfi yükü neytrallaşdıran kationlar dəyişdikdə, bu bölmələr Z formasına çevrilə bilər, DNT-nin digər bölmələri isə zəncir klassik B formasında qalır. Belə bir keçidin mümkünlüyü onu göstərir ki, DNT cüt spiralındakı iki zəncir dinamik vəziyyətdədir və bir-birinə nisbətən açıla bilər, sağ əlli formadan sol əlliyə və əksinə keçə bilər. DNT strukturunun konformasiya çevrilməsinə imkan verən belə labilliyin bioloji nəticələri hələ tam başa düşülməyib. Z-DNT bölmələrinin müəyyən genlərin ifadəsini tənzimləməkdə müəyyən rol oynadığına və genetik rekombinasiyada iştirak etdiyinə inanılır.
DNT-nin Z forması, fosfodiester onurğasının molekulun oxu boyunca ziqzaq şəklində yerləşdiyi sol əlli ikiqat spiraldır. Beləliklə, molekulun adı (ziqzaq)-DNK. Z-DNT təbiətdə məlum olan ən az bükülmüş (hər növbədə 12 əsas cüt) və ən nazik DNT-dir. Qonşu nukleotidlər arasındakı məsafə 0,38 nm, növbənin uzunluğu 4,56 nm, Z-DNT-nin diametri isə 1,8 nm-dir. Bundan əlavə, bu DNT molekulunun görünüşü tək bir oluğun olması ilə fərqlənir.
DNT-nin Z forması prokaryotik və eukaryotik hüceyrələrdə aşkar edilmişdir. İndi Z-formasını DNT-nin B-formasından fərqləndirə bilən antikorlar əldə edilmişdir. Bu antikorlar Drosophila (Dr. melanogaster) tüpürcək vəzi hüceyrələrinin nəhəng xromosomlarının müəyyən bölgələrinə bağlanır. Bu xromosomların qeyri-adi quruluşuna görə bağlanma reaksiyasını izləmək asandır, burada sıx bölgələr (disklər) daha az sıx bölgələrlə (disklər arası) ziddiyyət təşkil edir. Z-DNT bölgələri interdisklərdə yerləşir. Buradan belə nəticə çıxır ki, Z formasının ayrı-ayrı hissələrinin ölçüləri hələ də məlum olmasa da, əslində təbii şəraitdə mövcuddur.

(inverterlər) DNT-də ən məşhur və tez-tez rast gəlinən əsas ardıcıllıqlardır. Palindrom soldan sağa və əksinə oxunan söz və ya ifadədir. Belə söz və ya ifadələrə misal olaraq: DAXMA, KAZAK, DAŞQIN VƏ AZORUN PƏNƏNİNƏ DÜŞMÜŞ GÜL. DNT bölmələrinə tətbiq olunduqda bu termin (palindrom) zəncir boyu nukleotidlərin sağdan sola və soldan sağa eyni növbələşməsini ifadə edir (“daxma” sözündəki hərflər və s. kimi).
Palindrom iki DNT zəncirinə nisbətən ikinci dərəcəli simmetriyaya malik olan əsas ardıcıllığın tərs təkrarlarının olması ilə xarakterizə olunur. Bu cür ardıcıllıqlar, aşkar səbəblərə görə, özünü tamamlayır və saç tıxacları və ya xaç formalı strukturlar yaratmağa meyllidirlər (şəkil). Saç sancaqları tənzimləyici zülallara xromosom DNT-nin genetik mətninin haradan kopyalandığını tanımağa kömək edir.
Eyni DNT zəncirində ters çevrilmiş təkrar mövcud olduqda, ardıcıllığa güzgü təkrarı deyilir. Güzgü təkrarları özünü tamamlayıcı xüsusiyyətlərə malik deyil və buna görə də saç tıxacları və ya xaç formalı strukturlar yaratmaq iqtidarında deyil. Bu tip ardıcıllıqlar demək olar ki, bütün böyük DNT molekullarında olur və bir neçə baza cütündən bir neçə min əsas cütünə qədər dəyişə bilər.
Eukaryotik hüceyrələrdə xaçşəkilli strukturlar şəklində palindromların olması sübut edilməmişdir, baxmayaraq ki, E. coli hüceyrələrində in vivo olaraq müəyyən sayda xaç formalı strukturlar aşkar edilmişdir. RNT və ya tək zəncirli DNT-də özünü tamamlayan ardıcıllığın olması məhlullarda nuklein turşusu zəncirinin çoxlu “saç tıxaclarının” əmələ gəlməsi ilə xarakterizə olunan müəyyən bir məkan quruluşuna bükülməsinin əsas səbəbidir.

H-formalı DNTüç DNT zəncirindən əmələ gələn spiraldır - DNT üçlü spiral. Bu, Hoogsteen cütünü meydana gətirərək, əsas yivinə uyğunlaşan üçüncü tək zəncirli DNT zəncirinə malik Watson-Crick qoşa spiral kompleksidir.
Belə bir tripleksin əmələ gəlməsi DNT-nin qoşa spiralının elə bükülməsi nəticəsində baş verir ki, onun bölməsinin yarısı qoşa spiral şəklində qalır, digər yarısı isə ayrılır. Bu halda, ayrılmış sarmallardan biri qoşa sarmalın birinci yarısı ilə yeni bir quruluş meydana gətirir - üçlü spiral, ikincisi isə struktursuz, tək telli kəsik şəklində çıxır. Bu struktur keçidin bir xüsusiyyəti, protonları yeni quruluşu sabitləşdirən mühitin pH-dan kəskin asılılığıdır. Bu xüsusiyyətinə görə yeni struktur DNT-nin H forması adlanırdı, onun əmələ gəlməsi güzgü təkrarı olan homopurin-homopirimidin bölgələrini ehtiva edən super qıvrılmış plazmidlərdə aşkar edilmişdir.
Sonrakı tədqiqatlarda müəyyən edilmişdir ki, bəzi homopurin-homopirimidin ikiqat zəncirli polinükleotidlərin üç zəncirli strukturun formalaşması ilə struktur keçidini həyata keçirmək mümkündür:
- bir homopurin və iki homopirimidin teli ( Py-Pu-Py tripleksi) [Hoogsteen qarşılıqlı əlaqəsi].
Py-Pu-Py tripleksinin tərkib blokları kanonik izomorf CGC+ və TAT triadalarıdır. Tripleksin sabitləşməsi üçün CGC+ triadasının protonasiyası tələb olunur, ona görə də bu triplekslər məhlulun pH-dan asılıdır.
- bir homopirimidin və iki homopurin teli ( Py-Pu-Pu üçlüyü) [Ters Hoogsteen qarşılıqlı əlaqəsi].
Py-Pu-Pu tripleksinin tərkib blokları kanonik izomorf CGG və TAA triadalarıdır. Py-Pu-Pu triplekslərinin əsas xüsusiyyəti onların sabitliyinin ikiqat yüklü ionların mövcudluğundan asılılığıdır və müxtəlif ardıcıllığın triplekslərini sabitləşdirmək üçün müxtəlif ionlar tələb olunur. Py-Pu-Pu triplekslərinin əmələ gəlməsi onları təşkil edən nukleotidlərin protonasiyasını tələb etmədiyi üçün belə triplekslər neytral pH-da mövcud ola bilər.
Qeyd: birbaşa və tərs Hoogsteen qarşılıqlı təsirləri 1-metiltiminin simmetriyası ilə izah olunur: 180° fırlanma nəticəsində O2 atomu O4 atomunun yerini alır, hidrogen bağları sistemi isə qorunub saxlanılır.
İki növ üçlü sarmal məlumdur:
- Üçüncü ipin polaritesinin Watson-Crick dupleksinin homopurin zəncirinin polaritesi ilə üst-üstə düşdüyü paralel üçlü sarmallar
- üçüncü və homopurin zəncirlərinin qütblərinin əks olduğu antiparalel üçlü sarmallar.

G-dördlük- 4 zəncirli DNT. Bu quruluş, dörd guaninin sözdə G-quadrupleksi - dörd guaninin dəyirmi rəqsini meydana gətirən dörd guanin olduqda formalaşır.
Bu cür strukturların yaranma ehtimalına dair ilk göstərişlər Watson və Crick-in sıçrayış işlərindən çox əvvəl - 1910-cu ildə alındı. Sonra alman kimyaçısı İvar Banq aşkar etdi ki, DNT-nin tərkib hissələrindən biri - quanosinik turşu yüksək konsentrasiyalarda gel əmələ gətirir, DNT-nin digər komponentlərində isə bu xüsusiyyət yoxdur.
1962-ci ildə rentgen şüalarının difraksiya metodundan istifadə etməklə bu gelin hüceyrə quruluşunu qurmaq mümkün olmuşdur. Bir-birini bir dairədə birləşdirən və xarakterik bir kvadrat meydana gətirən dörd guanin qalığından ibarət olduğu ortaya çıxdı. Mərkəzdə əlaqə bir metal ionu (Na, K, Mg) tərəfindən dəstəklənir. DNT-də çoxlu guanin varsa, eyni strukturlar meydana gələ bilər. Bu düz kvadratlar (G-kvartetlər) kifayət qədər sabit, sıx strukturlar (G-dördlüklər) yaratmaq üçün yığılmışdır.
DNT-nin dörd ayrı zəncirindən dörd zəncirli komplekslər toxuna bilər, lakin bu, bir istisnadır. Daha tez-tez bir nuklein turşusu sadəcə olaraq bir düyünə bağlanır, xarakterik qalınlaşmalar əmələ gətirir (məsələn, xromosomların uclarında) və ya bəzi guaninlə zəngin bölgədə ikiqat zəncirli DNT yerli dördbucaqlı meydana gətirir.
Xromosomların uclarında - telomerlərdə və şiş promotorlarında dördlüklərin olması ən çox öyrənilmişdir. Bununla belə, insan xromosomlarında bu cür DNT-nin lokalizasiyasının tam mənzərəsi hələ də məlum deyil.
Xətti formada olan bu qeyri-adi DNT strukturlarının hamısı B-formalı DNT ilə müqayisədə qeyri-sabitdir. Bununla belə, DNT çox vaxt topoloji gərginliyin dairəvi formasında super qıvrılma adlanan şeyə malik olduqda mövcuddur. Bu şəraitdə qeyri-kanonik DNT strukturları asanlıqla formalaşır: Z-formaları, “xaçlar” və “saç sancaqları”, H-formaları, guanin kvadrupleksləri və i-motiflər.
- Supercoiled forma - pentoza fosfat onurğasına zərər vermədən hüceyrə nüvəsindən ayrıldıqda qeyd olunur. Çox bükülmüş qapalı üzüklər formasına malikdir. Həddindən artıq bükülmüş vəziyyətdə, DNT ikiqat sarmal ən azı bir dəfə "özünə bükülür", yəni ən azı bir superturn ehtiva edir (səkkiz rəqəm şəklini alır).
- DNT-nin rahat vəziyyəti - tək qırılma ilə müşahidə olunur (bir zəncirinin qırılması). Bu zaman super sarmallar yox olur və DNT qapalı halqa şəklini alır.
- DNT-nin xətti forması qoşa spiralın iki zəncirinin qırıldığı zaman müşahidə olunur.
DNT-nin üçüncü quruluşu
DNT-nin üçüncü quruluşu qoşa spiral molekulun fəzada əlavə burulması - onun superburulması nəticəsində əmələ gəlir. Eukaryotik hüceyrələrdə DNT molekulunun super qıvrılması prokariotlardan fərqli olaraq zülallarla komplekslər şəklində baş verir.
Eukariotların demək olar ki, bütün DNT-si nüvələrin xromosomlarında olur, yalnız kiçik bir hissəsi mitoxondrilərdə, bitkilərdə isə plastidlərdə olur. Eukaryotik hüceyrələrin (insan xromosomları da daxil olmaqla) xromosomlarının əsas maddəsi ikiqat zəncirli DNT, histon və qeyri-histon zülallarından ibarət xromatindir.
Histon xromatin zülalları
Histonlar xromatinin 50%-ni təşkil edən sadə zülallardır. Bütün tədqiq edilmiş heyvan və bitki hüceyrələrində histonların beş əsas sinfi aşkar edilmişdir: H1, H2A, H2B, H3, H4, ölçüləri, amin turşularının tərkibi və yükü ilə fərqlənir (həmişə müsbət).
Məməli histonu H1 təxminən 215 amin turşusu olan tək polipeptid zəncirindən ibarətdir; digər histonların ölçüləri 100 ilə 135 amin turşusu arasında dəyişir. Onların hamısı spiral halındadır və diametri təxminən 2,5 nm olan qlobula bükülür və qeyri-adi dərəcədə böyük miqdarda müsbət yüklü amin turşuları lizin və arginin ehtiva edir. Histonlar asetilləşə, metilləşə, fosforilləşə, poli(ADP)-ribosilləşə bilər və H2A və H2B histonları ubiquitinə kovalent şəkildə bağlıdır. Histonların strukturunun formalaşmasında və funksiyaların yerinə yetirilməsində bu cür modifikasiyaların rolu hələ tam aydınlaşdırılmamışdır. Güman edilir ki, bu, onların DNT ilə qarşılıqlı əlaqədə olmaq və gen fəaliyyətini tənzimləmək mexanizmlərindən birini təmin etmək qabiliyyətidir.
Histonlar DNT ilə ilk növbədə DNT-nin mənfi yüklü fosfat qrupları və histonların müsbət yüklü lizin və arginin qalıqları arasında əmələ gələn ion bağları (duz körpüləri) vasitəsilə qarşılıqlı əlaqədə olurlar.
Qeyri-histon xromatin zülalları
Qeyri-histon zülalları, histonlardan fərqli olaraq, çox müxtəlifdir. DNT-ni bağlayan qeyri-histon zülallarının 590-a qədər müxtəlif fraksiyaları təcrid edilmişdir. Onlara turşu zülallar da deyilir, çünki strukturlarında turşu amin turşuları üstünlük təşkil edir (onlar polianionlardır). Qeyri-histon zülallarının müxtəlifliyi xromatin fəaliyyətinin xüsusi tənzimlənməsi ilə bağlıdır. Məsələn, DNT replikasiyası və ifadəsi üçün lazım olan fermentlər müvəqqəti olaraq xromatinə bağlana bilər. Digər zülallar, məsələn, müxtəlif tənzimləmə proseslərində iştirak edənlər, DNT ilə yalnız xüsusi toxumalarda və ya müəyyən diferensiallaşma mərhələlərində bağlanırlar. Hər bir zülal DNT nukleotidlərinin müəyyən bir ardıcıllığını tamamlayır (DNT sahəsi). Bu qrupa daxildir:
- sahəyə xas sink barmaq zülalları ailəsi. Hər bir "sink barmağı" 5 nukleotid cütündən ibarət xüsusi bir yeri tanıyır.
- sahəyə məxsus zülallar ailəsi - homodimerlər. Belə bir zülalın DNT ilə təmasda olan fraqmenti spiral-dönmə-heliks quruluşuna malikdir.
- yüksək hərəkətli gel zülalları (HMG zülalları) daim xromatinlə əlaqəli olan struktur və tənzimləyici zülallar qrupudur. Onların molekulyar çəkisi 30 kDa-dan azdır və yüklənmiş amin turşularının yüksək tərkibi ilə xarakterizə olunur. Aşağı molekulyar çəkiyə görə HMG zülalları poliakrilamid gel elektroforezi zamanı yüksək hərəkətliliyə malikdir.
- replikasiya, transkripsiya və təmir fermentləri.
DNT və RNT-nin sintezində iştirak edən struktur, tənzimləyici zülalların və fermentlərin iştirakı ilə nukleosom ipi yüksək qatılaşdırılmış zülal və DNT kompleksinə çevrilir. Yaranan quruluş orijinal DNT molekulundan 10.000 dəfə qısadır.

Xromatin
Xromatin nüvə DNT və qeyri-üzvi maddələri olan zülallar kompleksidir. Xromatinin əsas hissəsi qeyri-aktivdir. Sıx qablaşdırılmış, qatılaşdırılmış DNT ehtiva edir. Bu heterokromatindir. Qeyri-ifadə edilmiş bölgələrdən ibarət konstitutiv, genetik cəhətdən aktiv olmayan xromatin (peyk DNT) və fakultativ - bir sıra nəsillərdə qeyri-aktiv, lakin müəyyən şərtlərdə ifadə edə bilən var.
Aktiv xromatin (euchromatin) kondenssizdir, yəni. daha az sıx şəkildə qablaşdırılır. Müxtəlif hüceyrələrdə onun tərkibi 2 ilə 11% arasında dəyişir. Beyin hüceyrələrində ən çox - 10-11%, qaraciyər hüceyrələrində - 3-4 və böyrək hüceyrələrində - 2-3% olur. Euchromatinin aktiv transkripsiyası qeyd olunur. Üstəlik, onun struktur təşkili müəyyən bir orqanizm növünə xas olan eyni genetik DNT məlumatının ixtisaslaşmış hüceyrələrdə fərqli şəkildə istifadə edilməsinə imkan verir.
Elektron mikroskopda xromatinin təsviri muncuqlara bənzəyir: təxminən 10 nm ölçülü sferik qalınlaşmalar, ip kimi körpülərlə ayrılır. Bu sferik qalınlaşmalara nukleosomlar deyilir. Nukleosom xromatinin struktur vahididir. Hər bir nukleosom hər bir nukleosom nüvəsinə 1,75 sola dönmə əmələ gətirmək üçün 146 bp superburulmuş DNT seqmentindən ibarətdir. Nükleosom nüvəsi H2A, H2B, H3 və H4 histonlarından, hər tipdən iki molekuldan ibarət olan histon oktameridir (şəkil 9), diametri 11 nm və qalınlığı 5,7 nm olan diskə bənzəyir. Beşinci histon, H1, nukleosom nüvəsinin bir hissəsi deyil və DNT-nin histon oktamerinə sarılması prosesində iştirak etmir. İkiqat spiralın nukleosom nüvəsinə daxil olduğu və çıxdığı yerlərdə DNT ilə əlaqə saxlayır. Bunlar interkor (bağlayıcı) DNT bölmələridir, uzunluğu hüceyrə tipindən asılı olaraq 40-50 nukleotid cütü arasında dəyişir. Nəticədə nukleosomlara daxil olan DNT fraqmentinin uzunluğu da dəyişir (186-196 nukleotid cütü).
Nukleosomlarda təxminən 90% DNT var, qalanları bağlayıcılardır. Nukleosomların "səssiz" xromatinin fraqmentləri olduğuna inanılır və bağlayıcı aktivdir. Bununla belə, nukleosomlar açılıb xətti ola bilər. Açılmış nukleosomlar artıq aktiv xromatindir. Bu, funksiyanın strukturdan asılılığını açıq şəkildə nümayiş etdirir. Güman etmək olar ki, qlobulyar nukleosomlarda nə qədər çox xromatin olarsa, bir o qədər az aktivdir. Aydındır ki, müxtəlif hüceyrələrdə istirahət xromatinin qeyri-bərabər nisbəti belə nukleosomların sayı ilə əlaqələndirilir.
Elektron mikroskopik fotoşəkillərdə, izolyasiya şəraitindən və uzanma dərəcəsindən asılı olaraq, xromatin nəinki qalınlaşması olan uzun iplik - nukleosomların "muncuqları" kimi, həm də diametrli daha qısa və sıx fibril (lif) kimi görünə bilər. 30 nm, onun formalaşması H1-in DNT və histonun bağlayıcı bölgəsinə bağlanması zamanı müşahidə olunur, bu da 30 nm diametrli bir solenoid meydana gətirmək üçün növbədə altı nukleosomun spiralının əlavə bükülməsinə səbəb olur. Bu zaman histon zülalı bir sıra genlərin transkripsiyasına mane ola bilər və bununla da onların fəaliyyətini tənzimləyə bilər.
DNT-nin yuxarıda təsvir edilən histonlarla qarşılıqlı təsiri nəticəsində orta diametri 2 nm və uzunluğu 57 nm olan 186 əsas cütdən ibarət DNT cüt spiralının seqmenti 10 nm diametrli bir spirala çevrilir. uzunluğu 5 nm. Bu spiral sonradan diametri 30 nm olan bir lifə sıxıldıqda, kondensasiya dərəcəsi daha altı dəfə artır.
Nəhayət, beş histonlu bir DNT dupleksinin qablaşdırılması DNT-nin 50 qat kondensasiyası ilə nəticələnir. Bununla belə, belə yüksək kondensasiya dərəcəsi belə metafaza xromosomunda DNT-nin demək olar ki, 50.000 - 100.000 qat sıxılmasını izah edə bilməz. Təəssüf ki, metafaza xromosomuna qədər sonrakı xromatinin qablaşdırılmasının təfərrüatları hələ məlum deyil, buna görə də biz yalnız bu prosesin ümumi xüsusiyyətlərini nəzərdən keçirə bilərik.
Xromosomlarda DNT-nin sıxılma səviyyələri
Hər bir DNT molekulu ayrı bir xromosomda qablaşdırılır. İnsan diploid hüceyrələrində hüceyrə nüvəsində yerləşən 46 xromosom var. Hüceyrədəki bütün xromosomların DNT-sinin ümumi uzunluğu 1,74 m-dir, lakin xromosomların qablaşdırıldığı nüvənin diametri milyonlarla dəfə kiçikdir. DNT-nin xromosomlarda və hüceyrə nüvəsindəki xromosomlarda belə yığcam qablaşdırılması DNT ilə müəyyən ardıcıllıqla qarşılıqlı əlaqədə olan müxtəlif histon və qeyri-histon zülalları ilə təmin edilir (yuxarıya bax). Xromosomlarda DNT-nin sıxlaşdırılması onun xətti ölçülərini təxminən 10.000 dəfə - təxminən 5 sm-dən 5 mikrona qədər azaltmağa imkan verir. Bir neçə səviyyəli sıxılma var (şək. 10).


- DNT ikiqat sarmal diametri 2 nm və uzunluğu bir neçə sm olan mənfi yüklü bir molekuldur.
- nukleosom səviyyəsi- xromatin elektron mikroskopda "muncuqlar" zənciri - nukleosomlar - "ip üzərində" görünür. Nukleosom həm euxromatində, həm də heterokromatində, interfaza nüvəsində və metafaza xromosomlarında olan universal struktur vahididir.
Nükleosomal sıxılma səviyyəsi xüsusi zülallar - histonlar tərəfindən təmin edilir. Səkkiz müsbət yüklü histon domeni mənfi yüklü DNT molekulunun sarıldığı nukleosomun nüvəsini təşkil edir. Bu, 7 dəfə qısalma verir, diametri isə 2-dən 11 nm-ə qədər artır.
- solenoid səviyyəsi
Xromosom təşkilatının solenoid səviyyəsi nukleosom filamentinin bükülməsi və diametri 20-35 nm olan daha qalın fibrillərin - solenoidlərin və ya superbidlərin meydana gəlməsi ilə xarakterizə olunur. Solenoid meydançası 11 nm-dir; hər növbədə təxminən 6-10 nukleosom var. Solenoid qablaşdırma superbid qablaşdırmadan daha çox hesab olunur, buna görə 20-35 nm diametrli bir xromatin fibril hər biri səkkiz nukleosomdan ibarət olan qranullar zənciri və ya superbidlərdir. Solenoid səviyyəsində DNT-nin xətti ölçüsü 6-10 dəfə azalır, diametri 30 nm-ə qədər artır.
- döngə səviyyəsi
Döngə səviyyəsi, təxminən 30-300 kb-lik döngələr əmələ gətirən, xüsusi DNT ardıcıllığını tanıyan və bağlayan qeyri-histon sahəsinə aid DNT-ni bağlayan zülallar tərəfindən təmin edilir. Döngə gen ifadəsini təmin edir, yəni. döngə yalnız struktur deyil, həm də funksional bir formalaşmadır. Bu səviyyədə qısalma 20-30 dəfə baş verir. Diametri 300 nm-ə qədər artır. Sitoloji preparatlarda amfibiya oositlərindəki “çıraq fırçaları” kimi ilgək formalı strukturları görmək olar. Bu döngələr superburuq kimi görünür və DNT domenlərini təmsil edir, ehtimal ki, transkripsiya və xromatin replikasiyası vahidlərinə uyğundur. Xüsusi zülallar döngələrin əsaslarını və bəlkə də bəzi daxili hissələrini düzəldir. Döngəyə bənzər domen təşkilatı metafaza xromosomlarında xromatinin daha yüksək dərəcəli spiral strukturlara qatlanmasını təşviq edir.
- domen səviyyəsi
Xromosom təşkilatının domen səviyyəsi kifayət qədər öyrənilməmişdir. Bu səviyyədə döngə domenlərinin əmələ gəlməsi qeyd olunur - 60% protein, 35% DNT və 5% RNT ehtiva edən 25-30 nm qalınlığında sapların (fibrillərin) strukturları hüceyrə dövrünün bütün fazalarında praktik olaraq görünməzdir. mitoz istisna olmaqla və hüceyrə nüvəsində bir qədər təsadüfi paylanır. Sitoloji preparatlarda amfibiya oositlərindəki “çıraq fırçaları” kimi ilgək formalı strukturları görmək olar.
Döngə domenləri öz bazasında nüvədaxili zülal matrisinə daxili qoşma saytları adlanan yerlərə yapışdırılır, tez-tez MAR/SAR ardıcıllığı kimi istinad edilir (MAR, İngilis matrisi ilə əlaqəli bölgədən; SAR, İngilis iskele əlavə bölgələrindən) - A/T nukleotid cütlərinin yüksək məzmunu (>65%) ilə xarakterizə olunan bir neçə yüz uzunluqlu əsas cütlük DNT fraqmentləri. Hər bir domen təkrarlanmanın tək mənşəyinə malikdir və avtonom superhelikal vahid kimi fəaliyyət göstərir. İstənilən döngə domenində çoxlu transkripsiya vahidləri var, onların işləməsi çox güman ki, əlaqələndirilir - bütün domen ya aktiv və ya qeyri-aktiv vəziyyətdədir.
Domen səviyyəsində, ardıcıl xromatin qablaşdırması nəticəsində DNT-nin xətti ölçülərində təxminən 200 dəfə (700 nm) azalma baş verir.
- xromosom səviyyəsi
Xromosom səviyyəsində profaza xromosomunun metafaza xromosomuna kondensasiyası qeyri-histon zülallarının eksenel çərçivəsi ətrafında döngə domenlərinin sıxılması ilə baş verir. Bu super qıvrılma hüceyrədəki bütün H1 molekullarının fosforlaşması ilə müşayiət olunur. Nəticədə, metafaza xromosomu sıx bir spiralə bükülmüş, sıx şəkildə yığılmış solenoid döngələr kimi təsvir edilə bilər. Tipik bir insan xromosomunda 2600-ə qədər döngə ola bilər. Belə bir quruluşun qalınlığı 1400 nm-ə (iki xromatid) çatır və DNT molekulu 104 dəfə qısaldılır, yəni. 5 sm uzanan DNT-dən 5 µm-ə qədər.
Xromosomların funksiyaları
Ekstraxromosom mexanizmləri ilə qarşılıqlı əlaqədə xromosomlar təmin edir
- irsi məlumatların saxlanması
- mobil təşkilat yaratmaq və saxlamaq üçün bu məlumatdan istifadə
- irsi məlumatların oxunmasının tənzimlənməsi
- genetik materialın öz-özünə təkrarlanması
- genetik materialın ana hüceyrədən qız hüceyrələrə köçürülməsi.
Xromatinin bir bölgəsi aktivləşdirildikdə, yəni. transkripsiya zamanı ondan əvvəlcə histon H1, sonra isə histon okteti tərsinə çıxarılır. Bu, xromatinin dekondensasiyasına, 30 nm xromatin fibrilinin 10 nm fibrilə ardıcıl keçidinə və daha sonra sərbəst DNT bölmələrinə açılmasına səbəb olur, yəni. nukleosom strukturunun itirilməsi.
1953-cü ildə DNT kimi bir maddənin molekulyar təşkili prinsipinin kəşfindən sonra molekulyar biologiya inkişaf etməyə başladı. Daha sonra tədqiqat prosesində alimlər DNT-nin necə rekombinasiya olunduğunu, onun tərkibini və insan genomumuzun necə qurulduğunu öyrəndilər.
Hər gün molekulyar səviyyədə mürəkkəb proseslər baş verir. DNT molekulu necə qurulmuşdur, nədən ibarətdir? Bəs DNT molekulları hüceyrədə hansı rolu oynayır? Qoşa zəncir daxilində baş verən bütün proseslər haqqında ətraflı danışaq.
İrsi məlumat nədir?
Bəs hər şey haradan başladı? Hələ 1868-ci ildə onu bakteriyaların nüvələrində tapdılar. Və 1928-ci ildə N. Koltsov canlı orqanizm haqqında bütün genetik məlumatların DNT-də şifrələndiyi nəzəriyyəsini irəli sürdü. Sonra J. Watson və F. Crick 1953-cü ildə indi tanınmış DNT spiral üçün bir model tapdılar və bunun üçün layiqincə tanınma və mükafat aldılar - Nobel mükafatı.
Hər halda DNT nədir? Bu maddə 2 birləşmiş ipdən, daha doğrusu spiraldan ibarətdir. Belə bir zəncirin müəyyən məlumatlara malik olan hissəsinə gen deyilir.

DNT hansı zülalların hansı ardıcıllıqla əmələ gələcəyi ilə bağlı bütün məlumatları özündə saxlayır. DNT makromolekulu, fərdi kərpiclərin - nukleotidlərin ciddi ardıcıllığında qeydə alınan inanılmaz həcmli məlumatların maddi daşıyıcısıdır. Cəmi 4 nukleotid var, onlar kimyəvi və həndəsi cəhətdən bir-birini tamamlayır. Elmdə bu tamamlama və ya tamamlama prinsipi daha sonra təsvir ediləcəkdir. Bu qayda genetik məlumatın kodlaşdırılması və dekodlanmasında əsas rol oynayır.
DNT zəncirinin inanılmaz uzun olduğu üçün bu ardıcıllıqda təkrarlar yoxdur. Hər bir canlının özünəməxsus DNT zəncirinə malikdir.
DNT-nin funksiyaları
Funksiyalara irsi məlumatların saxlanması və onun nəslə ötürülməsi daxildir. Bu funksiya olmadan bir növün genomu min illər ərzində qorunub saxlanıla və inkişaf etdirilə bilməzdi. Ciddi gen mutasiyalarına məruz qalmış orqanizmlərin sağ qalma ehtimalı daha yüksəkdir və ya nəsil yaratmaq qabiliyyətini itirir. Növlərin degenerasiyasına qarşı təbii qorunma belə baş verir.

Başqa bir vacib funksiya saxlanılan məlumatın həyata keçirilməsidir. Hüceyrə ikiqat zəncirdə saxlanılan bu təlimatlar olmadan tək bir həyati zülal yarada bilməz.
Nuklein turşusu tərkibi
Nukleotidlərin özlərinin - DNT-nin tikinti bloklarının - nədən ibarət olduğu indi dəqiq məlumdur. Onların tərkibində 3 maddə var:
- Ortofosfor turşusu.
- Azotlu əsas. Pirimidin əsasları - yalnız bir üzük olan. Bunlara timin və sitozin daxildir. 2 üzük ehtiva edən purin əsasları. Bunlar guanin və adenindir.
- saxaroza. DNT-də deoksiriboza, RNT-də riboza var.
Nukleotidlərin sayı həmişə azotlu əsasların sayına bərabərdir. Xüsusi laboratoriyalarda nukleotid parçalanır və ondan azotlu əsas ayrılır. Bu nukleotidlərin fərdi xassələri və onlarda mümkün mutasiyalar belə öyrənilir.
İrsi informasiyanın təşkili səviyyələri
Təşkilatlanmanın 3 səviyyəsi var: genetik, xromosom və genomik. Yeni zülalın sintezi üçün lazım olan bütün məlumatlar zəncirin kiçik bir hissəsində - gendə olur. Yəni, gen informasiyanın kodlaşdırılmasının ən aşağı və ən sadə səviyyəsi hesab olunur.

Genlər də öz növbəsində xromosomlara yığılır. İrsi materialın daşıyıcısının bu təşkili sayəsində xüsusiyyət qrupları müəyyən qanunlara uyğun olaraq növbələşir və bir nəsildən digərinə ötürülür. Qeyd etmək lazımdır ki, orqanizmdə inanılmaz sayda gen var, lakin dəfələrlə rekombinasiya edildikdə belə məlumat itmir.
Bir neçə növ gen var:
- Funksional təyinatına görə 2 növ var: struktur və tənzimləyici ardıcıllıqlar;
- Hüceyrədə baş verən proseslərə təsirinə əsaslanaraq, fərqləndirirlər: supervital, öldürücü, şərti öldürücü genlər, həmçinin mutator və antimutator genlər.
Genlər xromosom boyunca xətti ardıcıllıqla düzülür. Xromosomlarda məlumat təsadüfi fokuslanmır, müəyyən bir nizam var. Hətta genlərin mövqelərini və ya lokuslarını göstərən bir xəritə də var. Məsələn, 18 nömrəli xromosomun uşağın gözlərinin rəngi haqqında məlumatları şifrələdiyi məlumdur.
Genom nədir? Bu, orqanizmin hüceyrəsindəki bütün nukleotid ardıcıllığına verilən addır. Genom fərdi deyil, bütöv bir növü xarakterizə edir.
İnsanın genetik kodu nədir?
Fakt budur ki, insan inkişafının bütün böyük potensialı artıq konsepsiya dövründədir. Ziqotun inkişafı və doğuşdan sonra uşağın böyüməsi üçün lazım olan bütün irsi məlumatlar genlərdə şifrələnir. DNT bölmələri irsi məlumatın ən əsas daşıyıcılarıdır.
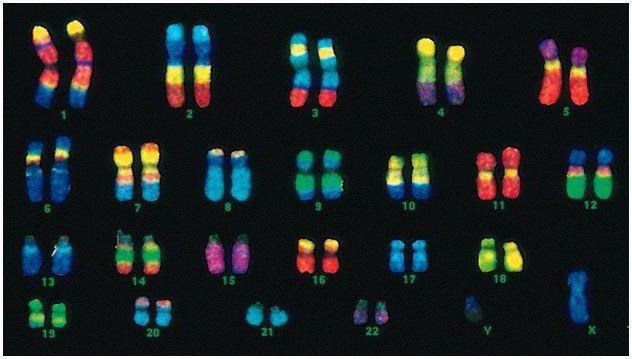
İnsanlarda 46 xromosom və ya 22 somatik cüt və hər bir valideyndən bir cinsi təyin edən xromosom var. Bu diploid xromosom dəsti insanın bütün fiziki görünüşünü, onun zehni və fiziki qabiliyyətlərini və xəstəliklərə qarşı həssaslığını kodlaşdırır. Somatik xromosomlar zahirən fərqlənmir, lakin onlar fərqli məlumat daşıyırlar, çünki onlardan biri atadan, digəri anadandır.
Kişi kodu qadın kodundan sonuncu cüt xromosomda - XY-də fərqlənir. Qadın diploid dəsti sonuncu cütdür, XX. Kişilər bioloji analarından bir X xromosomu alırlar və bu da qızlarına ötürülür. Cinsiyyət Y xromosomu oğullara ötürülür.
İnsan xromosomları ölçü baxımından çox dəyişir. Məsələn, xromosomların ən kiçik cütü 17 saylıdır. Ən böyük cüt isə 1 və 3-dür.
İnsanlarda qoşa spiralın diametri cəmi 2 nm-dir. DNT o qədər möhkəm bükülür ki, hüceyrənin kiçik nüvəsinə sığar, baxmayaraq ki, bükülməsə, uzunluğu 2 metrə qədər olardı. Sarmal uzunluğu yüz milyonlarla nukleotiddir.
Genetik kod necə ötürülür?
Beləliklə, DNT molekulları hüceyrə bölünməsində hansı rol oynayır? Genlər - irsi məlumatların daşıyıcıları - bədənin hər bir hüceyrəsində yerləşir. Bir çox canlılar öz kodlarını qız orqanizminə ötürmək üçün DNT-lərini 2 eyni spirala bölürlər. Buna replikasiya deyilir. Replikasiya prosesi zamanı DNT açılır və xüsusi “maşınlar” hər bir ipi tamamlayır. Genetik spiral ikiləşdikdən sonra nüvə və bütün orqanoidlər, sonra isə bütün hüceyrə bölünməyə başlayır.
Ancaq insanlarda fərqli bir gen ötürülməsi prosesi var - cinsi. Ata və ananın xüsusiyyətləri qarışıqdır, yeni genetik kodda hər iki valideynin məlumatları var.
İrsi məlumatların saxlanması və ötürülməsi DNT spiralının mürəkkəb təşkili sayəsində mümkündür. Axı dediyimiz kimi, zülalların strukturu genlərdə şifrələnir. Konsepsiya zamanı yaradıldıqdan sonra bu kod həyat boyu özünü kopyalayacaq. Orqan hüceyrələrinin yenilənməsi zamanı karyotip (xromosomların şəxsi dəsti) dəyişmir. Məlumatın ötürülməsi cinsi gametlərin - kişi və qadınların köməyi ilə həyata keçirilir.
Yalnız bir RNT zəncirini ehtiva edən viruslar öz məlumatlarını nəsillərinə ötürmək qabiliyyətinə malik deyillər. Buna görə də onların çoxalması üçün insan və ya heyvan hüceyrələri lazımdır.
İrsi məlumatların həyata keçirilməsi
Hüceyrə nüvəsində daim mühüm proseslər baş verir. Xromosomlarda qeydə alınan bütün məlumatlar amin turşularından zülal yaratmaq üçün istifadə olunur. Lakin DNT zənciri heç vaxt nüvəni tərk etmədiyi üçün başqa bir mühüm birləşmənin köməyinə ehtiyacı var: RNT. Nüvə membranına nüfuz edə və DNT zənciri ilə qarşılıqlı əlaqə qura bilən RNT-dir.
DNT və 3 növ RNT-nin qarşılıqlı təsiri ilə bütün kodlaşdırılmış məlumatlar həyata keçirilir. İrsi informasiyanın həyata keçirilməsi hansı səviyyədə baş verir? Bütün qarşılıqlı təsirlər nukleotid səviyyəsində baş verir. Messenger RNT DNT zəncirinin bir hissəsini kopyalayır və bu nüsxəni ribosoma gətirir. Burada nukleotidlərdən yeni molekulun sintezi başlayır.
mRNT-nin zəncirin lazımi hissəsini kopyalaya bilməsi üçün sarmal açılır və sonra yenidən kodlaşdırma prosesi başa çatdıqdan sonra yenidən bərpa olunur. Üstəlik, bu proses 1 xromosomun 2 tərəfində eyni vaxtda baş verə bilər.
Tamamlayıcılıq prinsipi
Onlar 4 nukleotiddən ibarətdir - adenin (A), guanin (G), sitozin (C), timin (T). Onlar tamamlayıcılıq qaydasına uyğun olaraq hidrogen bağları ilə bağlanır. E.Çarqaffın işi bu qaydanı yaratmağa kömək etdi, çünki alim bu maddələrin davranışında bəzi nümunələri qeyd etdi. E.Çarqaff kəşf etdi ki, adeninin timinlə molyar nisbəti birə bərabərdir. Və eyni şəkildə, guaninin sitozinə nisbəti həmişə birə bərabərdir.
Onun işinə əsaslanaraq, genetiklər nukleotidlərin qarşılıqlı əlaqəsi üçün bir qayda formalaşdırdılar. Komplementarlıq qaydası adeninin yalnız timinlə, quaninin isə yalnız sitozinlə birləşdiyini bildirir. Sarmalın dekodlanması və ribosomda yeni bir zülalın sintezi zamanı bu alternativ qayda transfer RNT-yə bağlanmış lazımi amin turşusunu tez tapmağa kömək edir.
RNT və onun növləri
İrsi məlumat nədir? DNT-nin ikiqat zəncirindəki nukleotidlər. RNT nədir? Onun işi nədir? RNT və ya ribonuklein turşusu DNT-dən məlumat çıxarmağa, onu deşifrə etməyə və tamamlayıcılıq prinsipinə əsaslanaraq hüceyrələr üçün lazım olan zülalları yaratmağa kömək edir.
Ümumilikdə 3 növ RNT var. Onların hər biri ciddi şəkildə öz funksiyasını yerinə yetirir.
- Məlumat (mRNT), ya da matrisa deyilir. O, birbaşa hüceyrənin mərkəzinə, nüvəyə daxil olur. Xromosomlardan birində zülal yaratmaq üçün lazım olan genetik materialı tapır və qoşa zəncirinin tərəflərindən birini kopyalayır. Kopyalama tamamlayıcılıq prinsipinə uyğun olaraq yenidən baş verir.
- Nəqliyyat bir tərəfində nukleotid dekoderləri, digər tərəfində isə əsas koda uyğun gələn amin turşuları olan kiçik molekuldur. tRNT-nin vəzifəsi onu "emalatxanaya", yəni lazımi amin turşusunu sintez etdiyi ribosoma çatdırmaqdır.
- rRNT ribosomdur.İstehsal olunan protein miqdarına nəzarət edir. 2 hissədən ibarətdir - amin turşusu və peptid bölməsi.
Deşifrənin yeganə fərqi RNT-də timin olmamasıdır. Burada timin əvəzinə urasil var. Lakin sonra, zülal sintezi zamanı tRNT hələ də bütün amin turşularını düzgün şəkildə quraşdırır. Məlumatın dekodlanmasında hər hansı bir uğursuzluq baş verərsə, mutasiya baş verir.
Zədələnmiş DNT molekulunun bərpası
Zədələnmiş ikiqat ipin bərpası prosesi təmir adlanır. Təmir prosesində zədələnmiş genlər çıxarılır.

Sonra elementlərin tələb olunan ardıcıllığı dəqiq şəkildə təkrarlanır və çıxarıldığı yerdən zəncirdə eyni yerə kəsilir. Bütün bunlar xüsusi kimyəvi maddələr - fermentlər sayəsində baş verir.
Niyə mutasiyalar baş verir?
Niyə bəzi genlər mutasiyaya başlayır və öz funksiyalarını yerinə yetirməyi dayandırır - həyati vacib irsi məlumatları saxlayır? Bu, dekodlaşdırma zamanı səhvə görə baş verir. Məsələn, adenin təsadüfən timinlə əvəz olunarsa.
Xromosom və genomik mutasiyalar da var. Xromosom mutasiyaları irsi məlumatların hissələri itirildikdə, çoxaldıqda və ya hətta köçürülüb başqa bir xromosoma daxil edildikdə baş verir.

Genomik mutasiyalar ən ciddidir. Onların səbəbi xromosomların sayında dəyişiklikdir. Yəni, bir cüt əvəzinə - diploid dəst, karyotipdə triploid dəst mövcuddur.
Triploid mutasiyaların ən məşhur nümunəsi Daun sindromudur ki, burada şəxsi xromosom dəsti 47-dir. Belə uşaqlarda 21-ci cütün yerində 3 xromosom əmələ gəlir.
Poliploidiya adlı məlum mutasiya da var. Lakin poliploidiya yalnız bitkilərdə olur.
DNT molekulu ikiqat sarmal meydana gətirən iki zəncirdən ibarətdir. Onun strukturu ilk dəfə 1953-cü ildə Francis Crick və James Watson tərəfindən deşifrə edilmişdir.
Əvvəlcə bir-birinin ətrafında bükülmüş bir cüt nukleotid zəncirindən ibarət DNT molekulu niyə bu xüsusi formaya sahib olduğu ilə bağlı suallar doğurdu. Alimlər bu fenomeni tamamlayıcılıq adlandırırlar, yəni onun zəncirlərində yalnız müəyyən nukleotidlər bir-birinin əksinə tapıla bilər. Məsələn, adenin həmişə timinlə, guanin isə həmişə sitozinin əksinədir. DNT molekulunun bu nukleotidləri tamamlayıcı adlanır.
Sxematik olaraq belə təsvir edilmişdir:
T - Ə
C - G
Bu cütlər amin turşularının sırasını təyin edən kimyəvi nukleotid bağı əmələ gətirir. Birinci halda, bir az zəifdir. C və G arasında əlaqə daha güclüdür. Tamamlayıcı olmayan nukleotidlər bir-biri ilə cüt əmələ gəlmir.
Bina haqqında
Deməli, DNT molekulunun quruluşu xüsusidir. O, bir səbəbdən bu formaya malikdir: fakt budur ki, nukleotidlərin sayı çox böyükdür və uzun zəncirləri yerləşdirmək üçün çox yer lazımdır. Məhz bu səbəbdən zəncirlər spiral bükülmə ilə xarakterizə olunur. Bu fenomen spirallaşma adlanır, ipləri təxminən beş-altı dəfə qısaltmağa imkan verir.
Bədən bu tip bəzi molekulları çox aktiv şəkildə istifadə edir, digərləri nadir hallarda. Sonuncu, spirallaşma ilə yanaşı, superspirallaşma kimi "yığcam qablaşdırma" da keçir. Və sonra DNT molekulunun uzunluğu 25-30 dəfə azalır.

Molekulun “qablaşdırılması” nədir?
Supercoiling prosesi histon zülallarını əhatə edir. Onlar iplik və ya çubuqun quruluşuna və görünüşünə malikdirlər. Onların üzərinə spiral iplər sarılır, onlar dərhal "kompakt şəkildə qablaşdırılır" və az yer tutur. Bu və ya digər ipdən istifadə etmək zərurəti yarandıqda, o, bir makaradan, məsələn, bir histon zülalından açılır və spiral iki paralel zəncirdə açılır. DNT molekulu bu vəziyyətdə olduqda, ondan lazımi genetik məlumatlar oxuna bilər. Bununla belə, bir şərt var. Məlumat əldə etmək yalnız DNT molekulunun strukturunun bükülməmiş formasına malik olduqda mümkündür. Oxumaq üçün əlçatan olan xromosomlar euxromatinlər adlanır və əgər onlar həddindən artıq bükülürsə, deməli onlar artıq heterokromatinlərdir.
Nuklein turşuları
Nuklein turşuları, zülallar kimi, biopolimerlərdir. Əsas funksiya irsi (genetik məlumatların) saxlanması, həyata keçirilməsi və ötürülməsidir. Onlar iki növdə olur: DNT və RNT (deoksiribonukleik və ribonukleik). Onların tərkibindəki monomerlər nukleotidlərdir, hər birində fosfor turşusu qalığı, beş karbonlu şəkər (dezoksiriboza/riboza) və azotlu əsas var. DNT koduna 4 növ nukleotid daxildir - adenin (A) / guanin (G) / sitozin (C) / timin (T). Onlar tərkibindəki azotlu əsasda fərqlənirlər.
DNT molekulunda nukleotidlərin sayı çox ola bilər - bir neçə mindən on və yüz milyonlara qədər. Belə nəhəng molekulları elektron mikroskop vasitəsilə tədqiq etmək olar. Bu vəziyyətdə, bir-birinə nukleotidlərin azotlu əsaslarının hidrogen bağları ilə bağlanan ikiqat polinukleotid zəncirlərini görə biləcəksiniz.

Araşdırma
Tədqiqat zamanı elm adamları DNT molekullarının növlərinin müxtəlif canlı orqanizmlərdə fərqli olduğunu aşkar etdilər. Həmçinin müəyyən edilmişdir ki, bir zəncirin guanini yalnız sitozinə, timin isə adeninə bağlana bilir. Bir zəncirdə nukleotidlərin düzülüşü paralel zəncirlə ciddi şəkildə uyğundur. Polinükleotidlərin bu tamamlayıcılığı sayəsində DNT molekulu ikiqat artırma və özünü çoxaltma qabiliyyətinə malikdir. Ancaq əvvəlcə, qoşalaşmış nukleotidləri məhv edən xüsusi fermentlərin təsiri altında tamamlayıcı zəncirlər ayrılır və sonra onların hər birində itkin zəncirin sintezi başlayır. Bu, hər bir hüceyrədə böyük miqdarda mövcud olan sərbəst nukleotidlərə görə baş verir. Bunun nəticəsində “ana molekul” əvəzinə tərkibi və quruluşu ilə eyni olan iki “qız” əmələ gəlir və DNT kodu orijinal olur. Bu proses hüceyrə bölünməsinin xəbərçisidir. Bütün irsi məlumatların ana hüceyrələrdən qız hüceyrələrinə, eləcə də bütün sonrakı nəsillərə ötürülməsini təmin edir.
Gen kodu necə oxunur?
Bu gün təkcə DNT molekulunun kütləsi hesablanmır, həm də əvvəllər elm adamları üçün əlçatmaz olan daha mürəkkəb məlumatları tapmaq mümkündür. Məsələn, bir orqanizmin öz hüceyrəsindən necə istifadə etməsi haqqında məlumatları oxuya bilərsiniz. Əlbəttə ki, əvvəlcə bu məlumat kodlaşdırılmış formadadır və müəyyən bir matris formasına malikdir və buna görə də RNT olan xüsusi bir daşıyıcıya daşınmalıdır. Ribonuklein turşusu nüvə membranı vasitəsilə hüceyrəyə nüfuz edə və içəridəki kodlanmış məlumatları oxuya bilir. Beləliklə, RNT nüvədən hüceyrəyə gizli məlumatların daşıyıcısıdır və DNT-dən onun tərkibində dezoksiriboza əvəzinə riboza, timin əvəzinə urasil olması ilə fərqlənir. Bundan əlavə, RNT tək zəncirlidir.

RNT sintezi
DNT-nin dərin təhlili göstərdi ki, RNT nüvəni tərk etdikdən sonra sitoplazmaya daxil olur və burada matris kimi ribosomlara (xüsusi ferment sistemləri) inteqrasiya oluna bilər. Alınan məlumatları rəhbər tutaraq, onlar protein amin turşularının müvafiq ardıcıllığını sintez edə bilirlər. Ribosom üçlü koddan əmələ gələn zülal zəncirinə hansı növ üzvi birləşmənin bağlanmalı olduğunu öyrənir. Hər bir amin turşusu onu kodlayan özünəməxsus üçlüyə malikdir.
Zəncirin formalaşması başa çatdıqdan sonra o, özünəməxsus məkan forması alır və öz hormonal, konstruktiv, fermentativ və digər funksiyalarını yerinə yetirə bilən zülala çevrilir. Hər hansı bir orqanizm üçün bu gen məhsuludur. Məhz ondan genlərin hər cür keyfiyyətləri, xassələri və təzahürləri müəyyən edilir.

Genlər
Ardıcıllıq prosesləri ilk növbədə bir DNT molekulunun strukturunda neçə genin olduğu haqqında məlumat əldə etmək üçün hazırlanmışdır. Və tədqiqatlar elm adamlarına bu məsələdə böyük irəliləyiş əldə etməyə imkan versə də, onların dəqiq sayını bilmək hələ mümkün deyil.
Cəmi bir neçə il əvvəl DNT molekullarında təxminən 100 min gen olduğu güman edilirdi. Bir az sonra bu rəqəm 80 minə qədər azaldı və 1998-ci ildə genetiklər bir DNT-də cəmi 50 min genin olduğunu bildirdilər ki, bu da ümumi DNT uzunluğunun yalnız 3%-ni təşkil edir. Lakin genetiklərin son nəticələri təəccüblü idi. İndi onlar genomda bu vahidlərin 25-40 mini olduğunu iddia edirlər. Məlum olub ki, xromosom DNT-nin yalnız 1,5%-i zülalların kodlaşdırılmasına cavabdehdir.
Araşdırma bununla da bitməyib. Paralel olaraq gen mühəndisliyi mütəxəssisləri qrupu müəyyən edib ki, bir molekuldakı genlərin sayı düz 32 mindir. Gördüyünüz kimi, hələ də qəti cavab almaq mümkün deyil. Çoxlu ziddiyyətlər var. Bütün tədqiqatçılar yalnız öz nəticələrinə etibar edirlər.
Təkamül var idi?
Molekulun təkamülünə dair heç bir dəlil olmamasına baxmayaraq (çünki DNT molekulunun quruluşu kövrək və kiçik ölçülüdür), elm adamları yenə də bir fərziyyə irəli sürdülər. Laboratoriya məlumatlarına əsaslanaraq, onlar aşağıdakı versiyanı səsləndirdilər: görünüşünün ilkin mərhələsində molekul qədim okeanlarda tapılan 32-yə qədər amin turşusunu ehtiva edən sadə özünü təkrarlayan peptid formasına sahib idi.
Öz-özünə çoxaldıqdan sonra təbii seçmə qüvvələri sayəsində molekullar özlərini xarici elementlərdən qorumaq qabiliyyəti əldə etdilər. Daha uzun yaşamağa və daha çox miqdarda çoxalmağa başladılar. Özlərini lipid qabarcığında tapan molekulların özünü çoxaltmaq üçün hər şansı var idi. Bir sıra ardıcıl dövrlər nəticəsində lipid baloncukları hüceyrə membranlarının formasını, sonra isə - tanınmış hissəciklər aldı. Qeyd etmək lazımdır ki, bu gün DNT molekulunun hər hansı bölməsi mürəkkəb və aydın fəaliyyət göstərən bir quruluşdur, onun bütün xüsusiyyətləri alimlər tərəfindən hələ tam öyrənilməmişdir.

Müasir dünya
Bu yaxınlarda İsrail alimləri saniyədə trilyonlarla əməliyyat yerinə yetirə bilən kompüter hazırlayıblar. Bu gün o, yer üzündəki ən sürətli avtomobildir. Bütün sirr ondan ibarətdir ki, innovativ qurğu DNT ilə işləyir. Professorlar deyirlər ki, yaxın gələcəkdə belə kompüterlər hətta enerji istehsal edə biləcək.
Bir il əvvəl Rehovotdakı (İsrail) Weizmann İnstitutunun mütəxəssisləri molekul və fermentlərdən ibarət proqramlaşdırıla bilən molekulyar hesablama maşınının yaradılması barədə məlumat vermişdilər. Silikon mikroçipləri onlarla əvəz etdilər. Bu günə qədər komanda daha da irəliləyiş əldə edib. İndi yalnız bir DNT molekulu kompüteri lazımi məlumatları və lazımi yanacaqla təmin edə bilər.
Biokimyəvi “nanokompüterlər” uydurma deyil, onlar artıq təbiətdə mövcuddur və hər bir canlıda təzahür edir. Ancaq çox vaxt insanlar tərəfindən idarə olunmur. İnsan, məsələn, “Pi” sayını hesablamaq üçün hələ heç bir bitkinin genomu üzərində əməliyyat apara bilməz.
Məlumatların saxlanması/emalı üçün DNT-dən istifadə ideyası ilk dəfə alimlərin ağlına 1994-cü ildə gəldi. Məhz o zaman sadə bir riyazi problemi həll etmək üçün molekuldan istifadə edildi. O vaxtdan bəri bir sıra tədqiqat qrupları DNT kompüterləri ilə bağlı müxtəlif layihələr təklif ediblər. Ancaq burada bütün cəhdlər yalnız enerji molekuluna əsaslanırdı. Belə bir kompüteri çılpaq gözlə görə bilməzsiniz, o, sınaq borusunda şəffaf suyun məhluluna bənzəyir. Orada heç bir mexaniki hissə yoxdur, ancaq trilyonlarla biomolekulyar qurğular var - və bu, yalnız bir damla mayenin içindədir!
İnsan DNT
İnsanlar 1953-cü ildə, elm adamları ilk dəfə dünyaya ikiqat zəncirli DNT modelini nümayiş etdirə bildikdə insan DNT-sinin növü ilə tanış oldular. Bunun üçün Kirk və Watson Nobel mükafatı aldılar, çünki bu kəşf 20-ci əsrdə əsas oldu.
Zaman keçdikcə, təbii ki, onlar sübut etdilər ki, strukturlaşdırılmış insan molekulu təkcə təklif olunan versiyadakı kimi görünə bilməz. Daha ətraflı DNT analizi apardıqdan sonra A-, B- və solaxay Z- formasını kəşf etdilər. Forma A- çox vaxt istisnadır, çünki o, yalnız nəmlik çatışmazlığı olduqda əmələ gəlir. Ancaq bu, yalnız laboratoriya tədqiqatlarında mümkündür; təbii mühit üçün bu, anormaldır, canlı hüceyrədə belə bir proses baş verə bilməz.
B forması klassikdir və ikiqat sağ əlli zəncir kimi tanınır, lakin Z forması yalnız sola əks istiqamətdə bükülmür, həm də daha ziqzaq görünüşünə malikdir. Alimlər həmçinin G-dördlük formasını da müəyyən ediblər. Onun strukturunda 2 yox, 4 sap var. Genetiklərin fikrincə, bu forma guaninin çox olduğu yerlərdə baş verir.

Süni DNT
Bu gün artıq süni DNT var ki, bu da həqiqi olanın eyni surətidir; təbii qoşa spiralın quruluşunu mükəmməl şəkildə izləyir. Ancaq orijinal polinükleotiddən fərqli olaraq, sünidə yalnız iki əlavə nukleotid var.
Dublyaj real DNT-nin müxtəlif tədqiqatlarından əldə edilən məlumatlar əsasında yaradıldığı üçün o, həm də kopyalana, özünü çoxalda və inkişaf edə bilər. Mütəxəssislər 20 ilə yaxındır ki, belə bir süni molekulun yaradılması üzərində işləyirlər. Nəticə, genetik kodu təbii DNT ilə eyni şəkildə istifadə edə bilən heyrətamiz bir ixtiradır.
Mövcud dörd azotlu əsasa genetiklər təbii əsasların kimyəvi modifikasiyası ilə yaradılmış iki əlavə əlavə etdilər. Təbii DNT-dən fərqli olaraq, süni DNT olduqca qısa olduğu ortaya çıxdı. O, yalnız 81 əsas cütdən ibarətdir. Bununla belə, o, həm də çoxalır və inkişaf edir.
Süni şəkildə əldə edilən molekulun təkrarlanması polimeraza zəncirvari reaksiya sayəsində baş verir, lakin indiyə qədər bu, müstəqil olaraq deyil, alimlərin müdaxiləsi ilə baş verir. Sözügedən DNT-yə müstəqil olaraq lazımi fermentləri əlavə edərək, xüsusi hazırlanmış maye mühitə yerləşdirirlər.
Yekun nəticə
DNT inkişafı prosesi və son nəticəsi mutasiyalar kimi müxtəlif amillərdən təsirlənə bilər. Bu, analiz nəticəsinin etibarlı və etibarlı olması üçün maddə nümunələrinin öyrənilməsini zəruri edir. Buna misal olaraq atalıq testini göstərmək olar. Lakin mutasiya kimi hadisələrin nadir olması bizi sevindirməyə bilməz. Buna baxmayaraq, analiz əsasında daha dəqiq məlumat əldə etmək üçün maddə nümunələri həmişə təkrar yoxlanılır.

Bitki DNT
Yüksək ardıcıllıq texnologiyaları (HTS) sayəsində genomika sahəsində bir inqilab edildi - bitkilərdən DNT çıxarılması da mümkündür. Təbii ki, bitki materialından yüksək keyfiyyətli molekulyar çəkidə DNT-nin alınması mitoxondriya və xloroplast DNT-nin çoxlu sayda nüsxəsi, həmçinin polisaxaridlərin və fenol birləşmələrinin yüksək səviyyədə olması səbəbindən müəyyən çətinliklər yaradır. Bu vəziyyətdə nəzərdən keçirdiyimiz strukturu təcrid etmək üçün müxtəlif üsullardan istifadə olunur.
DNT-də hidrogen bağı
DNT molekulundakı hidrogen bağı elektronmənfi atoma bağlanmış müsbət yüklü hidrogen atomu arasında yaranan elektromaqnit cazibəsindən məsuldur. Bu dipol qarşılıqlı təsir kimyəvi bağ meyarına cavab vermir. Ancaq molekullararası və ya molekulun müxtəlif hissələrində, yəni intramolekulyar şəkildə baş verə bilər.

Hidrogen atomu bağın donoru olan elektronmənfi atoma bağlanır. Elektromənfi atom azot, flüor və ya oksigen ola bilər. O, - mərkəzsizləşdirmə vasitəsilə - elektron buludunu hidrogen nüvəsindən özünə çəkir və hidrogen atomunu (qismən) müsbət yüklü edir. H-nin ölçüsü digər molekul və atomlarla müqayisədə kiçik olduğu üçün yükü də kiçikdir.
DNT-nin dekodlanması
DNT molekulunu deşifrə etməzdən əvvəl alimlər əvvəlcə çoxlu sayda hüceyrə götürürlər. Ən dəqiq və uğurlu iş üçün onlardan təxminən bir milyona ehtiyac var. Tədqiqat zamanı əldə edilən nəticələr daim müqayisə edilir və qeyd olunur. Bu gün genomun dekodlanması artıq nadir deyil, əlçatan bir prosedurdur.

Təbii ki, tək bir hüceyrənin genomunun deşifrə edilməsi praktiki olmayan bir məşqdir. Bu cür tədqiqatlar zamanı əldə edilən məlumatlar alimləri maraqlandırmır. Ancaq başa düşmək lazımdır ki, bütün mövcud dekodlaşdırma üsulları, mürəkkəbliyinə baxmayaraq, kifayət qədər təsirli deyil. Onlar yalnız DNT-nin 40-70%-ni oxumağa imkan verəcəklər.
Bununla belə, Harvard professorları bu yaxınlarda genomun 90%-nin deşifrə edilə biləcəyi metodu açıqladılar. Texnika təcrid olunmuş hüceyrələrə primer molekulların əlavə edilməsinə əsaslanır və onların köməyi ilə DNT replikasiyası başlayır. Ancaq hətta bu metodu uğurlu hesab etmək olmaz, elmdə açıq şəkildə istifadə edilməzdən əvvəl onu hələ də təkmilləşdirmək lazımdır.
