Molekul DNK je lociran. DNK i geni. Novi pogled na DNK
Biohemijske osnove nasljednosti.
Genetska uloga nukleinskih kiselina.
Nukleinske kiseline su biološki polimeri koji se nalaze u svim ćelijama, od primitivnih do složenih. Prvi ih je otkrio Johann Friedrich Miescher 1868. godine u stanicama bogatim nuklearnim materijalom (leukociti, sperma lososa). Termin "nukleinske kiseline" predložen je 1889.
Postoje dvije vrste nukleinskih kiselina: DNK, RNK (ATP - mononukleotid). DNK i RNK su molekuli - matrice. DNK se nalazi oko 6*10 -12 g u somatskim ćelijama: u jezgru, mitohondrijama. RNK je dio ribozoma i nalazi se u jezgru i citoplazmi.
Proučavanje i dokazivanje vodeće uloge nukleinskih kiselina u prijenosu nasljednih informacija provedeno je na virusnim česticama. Poznato je da je virus mozaika duhana virulentan na duhan i trputac. Virusna čestica se sastoji od 95% proteina i 5% nukleinske kiseline. Proteinski kapsid u virusnim česticama je zamijenjen, ali je nakon nekog vremena protein u oba soja transformiran u prethodni oblik.
Kod bakteriofaga koji inficiraju E. coli, proteini omotača faga su označeni radioaktivnim S, a DNK faga označena je radioaktivnim P. U bakterijskoj ćeliji inficiranoj fagom formirane su čestice faga koje su sadržavale samo radioaktivni P.
Struktura i funkcije molekula DNK i RNK.
Nukleinske kiseline su biopolimeri nepravilne strukture, čiji su monomeri nukleotidi. Nukleotid se sastoji od ostataka tri supstance: fosforne kiseline, ugljenih hidrata - pentoze, azotne baze. DNK nukleotidi sadrže ugljikohidrat deoksiribozu, a RNK ribozu. Ostaci azotnih baza purina i pirimidina koji čine DNK su adenin, gvanin, citozin, timin. Molekuli RNK sadrže adenin, guanin, citozin i uracil.
Nukleotidi su međusobno povezani preko ostatka fosforne kiseline jednog nukleotida i ugljikohidrata drugog jakom kovalentnom esterskom vezom, koja se naziva "kisenički most". Veza ide preko 5. atoma ugljikohidrata ugljikohidrata jednog nukleotida do 3. atoma ugljika ugljikohidrata drugog nukleotida. Nukleotidna sekvenca predstavlja primarnu strukturu nukleinskih kiselina. RNK je jedan polinukleotidni lanac. Struktura DNK je dvostruki polinukleotidni lanac, umotan u spiralu.
Sekundarna struktura DNK nastaje kada se pojavi drugi lanac DNK, raspoređen po principu komplementarnosti u odnosu na prvi. Drugi lanac je suprotan prvom (antiparalelan). Dušične baze leže u ravni koja je okomita na ravan molekula - ovo liči na spiralno stepenište. Ograde ovog stepeništa su ostaci fosforne kiseline i ugljikohidrata, a stepenice su dušične baze.
Dušične baze koje čine svaki nukleotid u suprotno usmerenim lancima sposobne su da formiraju komplementarne vodikove veze jedna s drugom (zbog postojećih funkcionalnih grupa u strukturi svake azotne baze). Adenil nukleotid je komplementaran timinu, guanil citozinu i obrnuto. Same po sebi, ove veze su krhke, ali molekul DNK koji je više puta "prošiven" cijelom dužinom takvim vezama predstavlja vrlo jaku vezu.
Komplementarnost- ovo je prostorno-strukturna i hemijska korespondencija azotnih baza jedna s drugom; pristaju jedna drugoj „kao ključ od brave“.
Jedan molekul DNK može sadržavati 10 8 ili više nukleotida.
Strukturu molekule DNK kao dvostruke antiparalelne spirale predložili su 1953. američki biolog James Watson i engleski fizičar Francis Crick.
Molekul DNK svakog živog organizma na planeti sastoji se od samo četiri tipa nukleotida, koji se međusobno razlikuju po dušičnim bazama koje sadrže: adenil, guanil, timin i citozin. U tome svestranost DNK. Njihov niz je drugačiji, a broj je beskonačan.
Za svaku vrstu živih organizama i za svaki organizam posebno, ovaj slijed je individualan i strogo specifično .
Posebnost Struktura DNK je da su hemijski aktivni delovi molekule - azotne baze, uronjeni u centar spirale i formiraju međusobno komplementarne veze, a ostaci dezoksiriboze i fosforne kiseline nalaze se na periferiji i pokrivaju pristup azotnim bazama. - hemijski su neaktivni. Ova struktura može zadržati hemijsku stabilnost dugo vremena. Šta je još potrebno za pohranjivanje nasljednih informacija? Upravo ove karakteristike strukture DNK određuju njenu sposobnost da kodira i reprodukuje genetske informacije.
Jaku strukturu DNK prilično je teško uništiti. Ipak, to se redovno dešava u ćeliji – tokom sinteze RNK i udvostručavanja samog molekula DNK pre deobe ćelije.
Duplikacija, DNK replikacija počinje činjenicom da poseban enzim - DNK polimeraza - odmotava dvostruku spiralu i razdvaja je u odvojene niti - formira se reduplikacijska viljuška. Enzim djeluje kao brava na patent zatvaraču. Na svakom jednolančanom lancu - ljepljivim krajevima reduplikacijske viljuške - sintetizira se novi lanac iz slobodnih nukleotida u karioplazmi prema principu komplementarnosti. U dva nova molekula DNK, jedan lanac ostaje izvorni lanac majke, a drugi ostaje novi kćerni lanac. Kao rezultat toga, umjesto jednog molekula DNK, pojavljuju se dva molekula sa istim tačnim nukleotidnim sastavom kao i originalni.
U živim sistemima susrećemo se s novom vrstom reakcija nepoznatih u neživoj prirodi. Zovu se reakcije sinteze matrice . Sinteza matrice je poput bacanja na matricu: novi molekuli se sintetiziraju u skladu sa planom koji je postavljen u strukturi postojećih molekula. Ove reakcije osiguravaju tačan slijed monomernih jedinica u sintetiziranim polimerima. Monomeri stižu na određenu lokaciju na molekulima koji služe kao matrica gdje se odvija reakcija. Ako bi se takve reakcije dogodile kao rezultat nasumičnih sudara molekula, one bi se odvijale beskonačno sporo. Sinteza složenih molekula na principu šablona se izvodi brzo i precizno uz pomoć enzima. Sinteza šablona je u osnovi najvažnijih reakcija u sintezi nukleinskih kiselina i proteina. Ulogu matriksa u ćeliji imaju molekuli nukleinskih kiselina DNK ili RNK. Monomerni molekuli iz kojih se sintetiše polimer - nukleotidi ili aminokiseline - u skladu sa principom komplementarnosti, nalaze se i fiksiraju na matrici po strogo definisanom redosledu. Zatim se monomerne jedinice povezuju u polimerni lanac, a gotov polimer napušta matricu. Nakon toga, matrica je spremna za sklapanje nove, potpuno iste polimerne molekule.
Reakcije matriksnog tipa su specifična karakteristika žive ćelije. Oni su osnova temeljnog svojstva svih živih bića - sposobnosti reprodukcije svoje vrste.
Funkcije nukleinskih kiselina– čuvanje i prenošenje nasljednih informacija. Molekuli DNK kodiraju informacije o primarnoj strukturi proteina. Sinteza mRNA molekula odvija se na matrici DNK. Ovaj proces se naziva "transkripcija". I-RNA u procesu “translacije” implementira informaciju u obliku niza aminokiselina u molekulu proteina.
DNK svake ćelije nosi informacije ne samo o strukturnim proteinima koji određuju oblik ćelije, već i o svim proteinima enzima, hormonskim proteinima i drugim proteinima, kao io strukturi svih vrsta RNK.
Možda nukleinske kiseline pružaju različite vrste biološke memorije – imunološke, neurološke itd., a također igraju značajnu ulogu u regulaciji biosintetskih procesa.
Povezane informacije.
Prvi dokaz uloge DNK kao nosioca nasljednih informacija u organizmima privukao je ogromnu pažnju proučavanju nukleinskih kiselina. Godine 1869. F. Miescher je izolovao posebnu supstancu iz ćelijskih jezgara, koju je nazvao nuklein. Nakon 20 godina ovaj naziv je zamijenjen terminom nukleinska kiselina. Godine 1924. R. Felgen je razvio metodu za citološko prepoznavanje nukleinskih kiselina kroz njihovo specifično bojenje i pokazao da je DNK lokalizirana u jezgrima stanica, a RNK u citoplazmi. Godine 1936. A.N. Belozersky i I.I. Dubrovskaya je izolirala DNK u čistom obliku iz jezgri biljnih stanica. Do početka 1930-ih. Razjašnjeni su osnovni hemijski principi strukture šećera nukleinskih kiselina, a 1953. godine stvoren je strukturni model DNK.
Osnovna strukturna jedinica nukleinskih kiselina je nukleotida, koji se sastoji od tri hemijski različita dela povezana kovalentnim vezama (slika 5.2).
Rice. 5.2. Strukturne formule: A- nukleotidi; b- DNK; V - RNA (vidi i str. 110)

Rice. 5.2. Kraj. Strukturne formule: A- nukleotidi; 6 - DNK; V- RNK
Prvi dio je šećer koji sadrži pet atoma ugljika: deoksiriboza u DNK i riboza u RNK.
Drugi dio nukleotida, purinska ili pirimidinska dušična baza, kovalentno je vezan za prvi atom ugljika šećera, formirajući strukturu tzv. nukleozid. DNK sadrži purinske baze - adenin(A) i guanin(D) - i pirimidinske baze - timina(T) i citozin(C). Odgovarajući nukleozidi se nazivaju deoksiadenozin, deoksiguanozin, deoksitimidin i deoksicitidin. RNK sadrži iste purinske baze kao i DNK, pirimidinska baza citozin, a umjesto timina sadrži uracil(U); odgovarajući nukleozidi se nazivaju adenozin, gvanozin, uridin i citidin.
Treći dio nukleotida je fosfatna grupa, koja povezuje susjedne nukleozide u polimerni lanac preko fosfodiestarskih veza između 5-ugljičnog atoma jednog šećera i 3-ugljičnog atoma drugog (slika 5.2, b, V). Nukleotidi nazivaju se nukleozidi s jednom ili više fosfatnih grupa vezanih esterskim vezama za 3" ili 5 ugljikovih atoma šećera. Sinteza nukleotida prethodi sintezi nukleinskih kiselina; shodno tome, nukleotidi su produkti kemijske ili enzimske hidrolize nukleinskih kiselina.
Nukleinske kiseline su vrlo dugi polimerni lanci koji se sastoje od mononukleotida povezanih 5- i 3'-fosfodiestarskim vezama. Intaktni molekul DNK sadrži, zavisno od vrste organizma, od nekoliko hiljada do mnogo miliona nukleotida, netaknut molekul RNK - od 100 do 100 hiljada ili više nukleotida.
Rezultati E. Chargaffove analize nukleotidnog sastava DNK različitih oblika vrsta pokazali su da molekularni odnos različitih azotnih baza - adenina, gvanina, timina, citozina - uveliko varira. Posljedično, dokazano je da DNK uopće nije monoton polimer koji se sastoji od identičnih tetranukleotida, kako se pretpostavljalo 40-ih godina. XX vijeka, te da u potpunosti posjeduje kompleksnost neophodnu za očuvanje i prenošenje nasljednih informacija u obliku specifične sekvence nukleotidnih baza.
Istraživanje E. Chargaffa također je otkrilo osobinu svojstvenu svim molekulima DNK: molarni sadržaj adenina jednak je sadržaju timina, a molarni sadržaj gvanina jednak je sadržaju citozina. Ove jednakosti se nazivaju Chargaffovo pravilo ekvivalencije: [A] = [T], [G] = [C]; broj purina je jednak broju pirimidina. Ovisno o vrsti, mijenja se samo omjer ([A] + [T])/([G] + [C]) (tabela 5.1).
|
Sastav baza |
Stav |
Asimetrija |
|||||
|
razlozi |
|||||||
|
(A + T)/(G + C) |
|||||||
|
Životinje |
|||||||
|
Kornjača |
|||||||
|
morski rak |
|||||||
|
Morski jež |
|||||||
|
Biljke, pečurke |
|||||||
|
pšenične klice |
|||||||
|
Mushroom Aspergillus niger |
|||||||
|
Bakterije |
|||||||
|
Escherichia coli |
|||||||
|
Staphylococcus aureus |
|||||||
|
Clostridium perfringens |
|||||||
|
Brucela abortus |
|||||||
|
Sarcina lutea |
|||||||
|
Bakteriofagi |
|||||||
|
FH 174 (virusni oblik) |
|||||||
|
FH 174 (replikacijski oblik) |
|||||||
Omjer baza se naziva nukleotidni koeficijent(vrsta) specifičnost. Chargaffovo otkriće formuliralo je važnu strukturnu osobinu DNK, koja se kasnije odrazila u strukturnom modelu DNK od strane J. Watsona i F. Cricka (1953), koji su zapravo pokazali da Chargaffova pravila ne nameću nikakva ograničenja na mogući broj kombinacija različite bazne sekvence sposobne da formiraju molekule DNK.
Koncept specifičnosti nukleotida formirao je osnovu za novu granu biologije - sistematika gena, koji funkcioniše tako što upoređuje sastav i strukturu nukleinskih kiselina kako bi izgradio prirodni sistem organizama.
Prema Watson-Crickovom modelu, molekula DNK se sastoji od dva polinukleotidna lanca (lanca, lanca), međusobno povezana poprečnim vodikovim vezama između azotnih baza prema komplementarnom principu (adenin jednog lanca je povezan sa dvije vodikove veze sa timin suprotnog lanca, a gvanin i citozin različitih lanaca su međusobno povezani trima vodoničnim vezama). U ovom slučaju, dva polinukleotidna lanca jednog molekula su antiparalelna, tj. nasuprot 3" kraja jednog lanca nalazi se 5" kraj drugog lanca i obrnuto (slika 5.3). Međutim, treba imati na umu savremene podatke da genetski materijal nekih virusa predstavljaju jednolančane (jednolančane) DNK molekule. Na osnovu podataka rendgenske difrakcije DNK, J. Watson i F. Crick su također zaključili da njegova dvolančana molekula ima sekundarnu strukturu u obliku spirale uvijene u smjeru s lijeva na desno, koja je kasnije nazvana 5 -forma (slika 5.4). Do danas je dokazano da je pored najčešćeg 5-forma moguće detektovati dijelove DNK koji imaju drugačiju konfiguraciju – kao desnoruki (forme A, C), i uvrnuti s desna na lijevo (lijevo uvijen, ili Z-oblik) (slika 5.4). Postoje određene razlike između ovih oblika sekundarne strukture DNK (tabela 5.2). Na primjer, udaljenost između dva susjedna para dušičnih baza u dvolančanoj spirali, izražena u nanometrima (nm), karakterizirana je različitim vrijednostima za 5-oblik i Z-oblik (0,34 i 0,38 nm, respektivno) . Na sl. Slika 5.5 prikazuje moderne trodimenzionalne modele "ljevorukih" i "desnorukih" oblika DNK.

Rice. 5.3. šematski prikaz primarne strukture fragmenta dvolančane DNK molekule: A - adenin; G - gvanin; T - timin; C - citozin

Rice. 5.4.

Tabela 5.2
Svojstva različitih oblika DNK dvostrukih spirala
Molekule RNK, ovisno o njihovim strukturnim i funkcionalnim karakteristikama, dijele se na nekoliko tipova: glasnička RNA (mRNA, ili mRNA), ribosomalna RNA (rRNA), transferna RNA (tRNA), mala nuklearna RNA (snRNA) itd. Molekuli DNK, RNK su uvijek jednolančani (jednolančani). Međutim, mogu formirati složenije (sekundarne) konfiguracije zbog komplementarnog povezivanja pojedinih sekcija takvog lanca na osnovu interakcije komplementarnih azotnih baza (A-U i G-C). Kao primjer, razmotrite konfiguraciju lista djeteline za molekulu RNK za prijenos fenilalanina (slika 5.6).

Rice. 5.6.
Godine 1953. D. Watson i F. Crick su predložili model strukture DNK, koji je bio zasnovan na sljedećim postulatima:
- 1. DNK je polimer koji se sastoji od nukleotida povezanih 3" i 5" fosfodiestarskim vezama.
- 2. Sastav DNK nukleotida je u skladu sa Chargaffovim pravilima.
- 3. Molekul DNK ima strukturu dvostruke spirale, koja podsjeća na spiralno stepenište, o čemu svjedoče uzorci difrakcije rendgenskih zraka lanaca DNK koji su prvi dobili M. Wilkins i R. Franklin.
- 4. Struktura polimera, prikazana kiselo-baznom titracijom nativne (prirodne) DNK, stabilizovana je vodoničnim vezama. Titracija i zagrijavanje nativne DNK uzrokuje primjetnu promjenu njenih fizičkih svojstava, posebno viskoznosti, pretvarajući je u denaturirani oblik, a kovalentne veze se ne uništavaju.
Nukleinske kiseline su visokomolekularne supstance koje se sastoje od mononukleotida, koji su međusobno povezani u polimerni lanac pomoću fosfodiestarskih veza od 3", 5" i na određeni način se pakuju u ćelijama.
Nukleinske kiseline su biopolimeri dva tipa: ribonukleinska kiselina (RNA) i deoksiribonukleinska kiselina (DNK). Svaki biopolimer sastoji se od nukleotida koji se razlikuju po ostatku ugljikohidrata (riboza, deoksiriboza) i jednoj od dušičnih baza (uracil, timin). Prema ovim razlikama, nukleinske kiseline su dobile svoje ime.
Struktura deoksiribonukleinske kiseline
Nukleinske kiseline imaju primarnu, sekundarnu i tercijarnu strukturu.
Primarna struktura DNK
Primarna struktura DNK je linearni polinukleotidni lanac u kojem su mononukleotidi povezani 3", 5" fosfodiestarskim vezama. Početni materijal za sklapanje lanca nukleinske kiseline u ćeliji je 5"-trifosfatni nukleozid, koji je, kao rezultat uklanjanja ostataka β i γ fosforne kiseline, sposoban da veže 3" atom ugljika drugog nukleozida. . Dakle, 3" atom ugljika jedne deoksiriboze je kovalentno vezan za atom ugljika od 5" druge deoksiriboze preko jednog ostatka fosforne kiseline i formira linearni polinukleotidni lanac nukleinske kiseline. Otuda i naziv: 3", 5" fosfodiestarske veze. Azotne baze ne učestvuju u povezivanju nukleotida jednog lanca (slika 1.).
Takva veza, između ostatka molekule fosforne kiseline jednog nukleotida i ugljikohidrata drugog, dovodi do formiranja pentozo-fosfatnog skeleta polinukleotidnog molekula, na kojem su azotne baze jedna za drugom pričvršćene na stranu. Njihov slijed rasporeda u lancima molekula nukleinskih kiselina striktno je specifičan za ćelije različitih organizama, tj. ima specifičan karakter (Chargaffovo pravilo).
Linearni DNK lanac, čija dužina zavisi od broja nukleotida uključenih u lanac, ima dva kraja: jedan se zove 3" kraj i sadrži slobodni hidroksil, a drugi se zove 5" kraj i sadrži fosforni kiseli ostatak. Krug je polaran i može imati smjer od 5"->3" i 3"->5". Izuzetak je kružna DNK.
Genetski "tekst" DNK sastoji se od kodnih "riječi" - trojki nukleotida zvanih kodoni. Dijelovi DNK koji sadrže informacije o primarnoj strukturi svih vrsta RNK nazivaju se strukturni geni.
Polinukleotidni DNK lanci dostižu gigantske veličine, pa se na određeni način pakuju u ćeliji.
Proučavajući sastav DNK, Chargaff (1949) je ustanovio važne obrasce u vezi sa sadržajem pojedinačnih baza DNK. Oni su pomogli da se otkrije sekundarna struktura DNK. Ovi obrasci se nazivaju Chargaffova pravila. Chargaff pravila
Ova pravila ukazuju na to da se prilikom konstruisanja DNK mora poštovati prilično stroga korespondencija (uparivanje) ne purinskih i pirimidinskih baza uopšte, već posebno timina sa adeninom i citozina sa gvaninom. Na osnovu ovih pravila, Watson i Crick su 1953. godine predložili model sekundarne strukture DNK, nazvan dvostruka spirala (sl.). |

Sekundarna struktura DNK
Sekundarna struktura DNK je dvostruka spirala, čiji su model predložili D. Watson i F. Crick 1953. godine.
Preduvjeti za izradu DNK modela
Kao rezultat početnih analiza, vjerovalo se da DNK bilo kojeg porijekla sadrži sva četiri nukleotida u jednakim molarnim količinama. Međutim, 1940-ih, E. Chargaff i njegove kolege, kao rezultat analize DNK izolovane iz raznih organizama, jasno su pokazali da sadrže azotne baze u različitim kvantitativnim omjerima. Chargaff je otkrio da iako su ovi omjeri isti za DNK iz svih ćelija iste vrste organizma, DNK različitih vrsta može se značajno razlikovati u sadržaju određenih nukleotida. Ovo sugerira da razlike u omjeru azotnih baza mogu biti povezane s nekom vrstom biološkog koda. Iako se pokazalo da je omjer pojedinačnih purinskih i pirimidinskih baza u različitim uzorcima DNK različit, pri upoređivanju rezultata testa pokazao se određeni obrazac: u svim uzorcima ukupan broj purina bio je jednak ukupnom broju pirimidina (A + G = T + C), količina adenina je bila jednaka količini timina (A = T), a količina gvanina je količina citozina (G = C). DNK izolirana iz stanica sisara općenito je bila bogatija adeninom i timinom i relativno siromašnija gvaninom i citozinom, dok je DNK iz bakterija bila bogatija gvaninom i citozinom i relativno siromašnija adeninom i timinom. Ovi podaci su činili važan dio činjeničnog materijala na osnovu kojeg je kasnije izgrađen Watson-Crick model strukture DNK.
Još jednu važnu indirektnu indikaciju moguće strukture DNK dali su podaci L. Paulinga o strukturi proteinskih molekula. Pauling je pokazao da je moguće nekoliko različitih stabilnih konfiguracija lanca aminokiselina u molekulu proteina. Jedna uobičajena konfiguracija peptidnog lanca, α-heliks, je pravilna spiralna struktura. S ovom strukturom moguće je stvaranje vodikovih veza između aminokiselina koje se nalaze na susjednim zavojima lanca. Pauling je opisao α-heličnu konfiguraciju polipeptidnog lanca 1950. godine i sugerirao da molekuli DNK vjerovatno imaju spiralnu strukturu koju drže vodonične veze.
Ipak, najvrednije informacije o strukturi molekule DNK dali su rezultati analize difrakcije rendgenskih zraka. X-zrake koje prolaze kroz DNK kristal podliježu difrakciji, odnosno odbijaju se u određenim smjerovima. Stepen i priroda skretanja zraka zavise od strukture samih molekula. Difrakcija rendgenskih zraka (slika 3) daje iskusnom oku brojne indirektne indikacije u vezi sa strukturom molekula supstance koja se proučava. Analiza uzoraka difrakcije rendgenskih zraka DNK dovela je do zaključka da su dušične baze (koje imaju ravan oblik) raspoređene kao hrpa ploča. Difrakcijski uzorci rendgenskih zraka otkrili su tri glavna perioda u strukturi kristalne DNK: 0,34, 2 i 3,4 nm.

Watson-Crick DNK model
Na osnovu Chargaffovih analitičkih podataka, Wilkinsovih rendgenskih uzoraka i istraživanja kemičara koji su pružili informacije o preciznim udaljenostima između atoma u molekuli, uglovima između veza datog atoma i veličini atoma, Watson i Crick je počeo da gradi fizičke modele pojedinačnih komponenti molekula DNK na određenoj skali i "prilagođava" ih jedni drugima na način da dobijeni sistem odgovara različitim eksperimentalnim podacima. [prikaži] .
Još ranije je bilo poznato da su susjedni nukleotidi u lancu DNK povezani fosfodiesterskim mostovima, povezujući 5"-ugljični atom deoksiriboze jednog nukleotida sa 3"-ugljični deoksiribozni atom sljedećeg nukleotida. Watson i Crick nisu sumnjali da period od 0,34 nm odgovara udaljenosti između uzastopnih nukleotida u lancu DNK. Nadalje, moglo bi se pretpostaviti da period od 2 nm odgovara debljini lanca. A da bi objasnili kojoj stvarnoj strukturi odgovara period od 3,4 nm, Watson i Crick, kao i Pauling ranije, sugerirali su da je lanac uvijen u obliku spirale (ili, tačnije, formira spiralnu liniju, jer spirala u strogom smislu ovih riječi se dobija kada zavojnice formiraju konusnu, a ne cilindričnu površinu u prostoru). Tada će period od 3,4 nm odgovarati udaljenosti između uzastopnih zavoja ove spirale. Takva spirala može biti vrlo gusta ili donekle rastegnuta, odnosno njeni zavoji mogu biti ravni ili strmi. Budući da je period od 3,4 nm tačno 10 puta veći od udaljenosti između uzastopnih nukleotida (0,34 nm), jasno je da svaki potpuni okret spirale sadrži 10 nukleotida. Iz ovih podataka, Watson i Crick su uspjeli izračunati gustinu polinukleotidnog lanca uvijenog u spiralu prečnika 2 nm, sa razmakom između zavoja od 3,4 nm. Ispostavilo se da bi takav lanac imao gustinu koja je upola manja od stvarne gustine DNK, što je već bilo poznato. Morao sam pretpostaviti da se molekul DNK sastoji od dva lanca – da je to dvostruka spirala nukleotida.
Sljedeći zadatak je, naravno, bio razjasniti prostorne odnose između dva lanca koji formiraju dvostruku spiralu. Nakon što su isprobali brojne opcije za raspored lanaca na svom fizičkom modelu, Watson i Crick su otkrili da se svi dostupni podaci najbolje podudaraju s opcijom u kojoj dvije polinukleotidne spirale idu u suprotnim smjerovima; u ovom slučaju, lanci koji se sastoje od ostataka šećera i fosfata formiraju površinu dvostruke spirale, a purini i pirimidini se nalaze unutra. Baze koje se nalaze jedna naspram druge, koje pripadaju dva lanca, povezane su u parovima vodoničnim vezama; Ove vodonične veze drže lance zajedno, fiksirajući tako ukupnu konfiguraciju molekula.
Dvostruka spirala DNK može se zamisliti kao ljestve od užeta koje su uvijene na spiralni način, tako da prečke ostaju horizontalne. Tada će dva uzdužna užeta odgovarati lancima ostataka šećera i fosfata, a prečke će odgovarati parovima dušičnih baza povezanih vodikovim vezama.
Kao rezultat daljeg proučavanja mogućih modela, Watson i Crick su zaključili da bi se svaka "prečka" trebala sastojati od jednog purina i jednog pirimidina; u periodu od 2 nm (što odgovara prečniku dvostruke spirale), ne bi bilo dovoljno prostora za dva purina, a dva pirimidina ne bi mogla biti dovoljno blizu jedan drugom da formiraju odgovarajuće vodonične veze. Detaljna studija detaljnog modela pokazala je da adenin i citozin, iako tvore kombinaciju odgovarajuće veličine, još uvijek ne mogu biti postavljeni na takav način da bi se između njih stvorile vodikove veze. Slični izvještaji natjerali su da se isključi kombinacija gvanin - timin, dok su se kombinacije adenin - timin i guanin - citozin pokazale sasvim prihvatljivim. Priroda vodikovih veza je takva da adenin formira par sa timinom, a gvanin sa citozinom. Ova ideja specifičnog uparivanja baza omogućila je da se objasni "Chargaffovo pravilo", prema kojem je u bilo kojoj molekuli DNK količina adenina uvijek jednaka sadržaju timina, a količina gvanina uvijek jednaka količini citozina. Dvije vodikove veze formiraju se između adenina i timina, a tri između gvanina i citozina. Zbog ove specifičnosti, formiranje vodikovih veza protiv svakog adenina u jednom lancu uzrokuje stvaranje timina na drugom; na isti način, samo citozin može biti nasuprot svakom guaninu. Dakle, lanci su komplementarni jedni drugima, odnosno sekvenca nukleotida u jednom lancu jedinstveno određuje njihov niz u drugom. Dva lanca idu u suprotnim smjerovima i njihove terminalne fosfatne grupe su na suprotnim krajevima dvostruke spirale.
Kao rezultat svog istraživanja, Watson i Crick su 1953. godine predložili model strukture molekula DNK (slika 3), koji je ostao relevantan do današnjih dana. Prema modelu, molekul DNK se sastoji od dva komplementarna polinukleotidna lanca. Svaki lanac DNK je polinukleotid koji se sastoji od nekoliko desetina hiljada nukleotida. U njemu susjedni nukleotidi formiraju pravilnu pentozo-fosfatnu kičmu zbog veze ostatka fosforne kiseline i deoksiriboze snažnom kovalentnom vezom. Dušične baze jednog polinukleotidnog lanca raspoređene su u strogo definisanom redosledu nasuprot azotnim bazama drugog. Izmjena dušičnih baza u polinukleotidnom lancu je nepravilna.
Raspored azotnih baza u lancu DNK je komplementaran (od grčkog "komplement" - dodavanje), tj. Timin (T) je uvijek protiv adenina (A), a samo citozin (C) je protiv guanina (G). Ovo se objašnjava činjenicom da A i T, kao i G i C, striktno odgovaraju jedno drugom, tj. dopunjuju jedno drugo. Ova korespondencija je određena hemijskom strukturom baza, koja omogućava stvaranje vodoničnih veza u purinskom i pirimidinskom paru. Postoje dvije veze između A i T, a tri između G i C. Ove veze obezbeđuju delimičnu stabilizaciju DNK molekula u svemiru. Stabilnost dvostruke spirale je direktno proporcionalna broju G≡C veza, koje su stabilnije u odnosu na A=T veze.
Poznata sekvenca rasporeda nukleotida u jednom lancu DNK omogućava da se, po principu komplementarnosti, uspostave nukleotidi drugog lanca.
Osim toga, utvrđeno je da se dušične baze koje imaju aromatičnu strukturu u vodenom rastvoru nalaze jedna iznad druge, formirajući, takoreći, hrpu novčića. Ovaj proces formiranja naslaga organskih molekula naziva se slaganje. Polinukleotidni lanci molekule DNK razmatranog Watson-Crickovog modela imaju slično fizičko-hemijsko stanje, njihove dušične baze su raspoređene u obliku hrpe novčića, između čijih ravnina nastaju van der Waalsove interakcije (interakcije slaganja).
Vodikove veze između komplementarnih baza (horizontalno) i interakcije slaganja između ravni baza u polinukleotidnom lancu zbog van der Waalsovih sila (vertikalno) pružaju molekuli DNK dodatnu stabilizaciju u prostoru.
Šećerno-fosfatne okosnice oba lanca okrenute su prema van, a baze okrenute prema unutra, jedna prema drugoj. Smjer lanaca u DNK je antiparalelan (jedan od njih ima smjer 5"->3, drugi - 3"->5", tj. kraj od 3" jednog lanca nalazi se nasuprot kraju od 5" drugi.). Lanci formiraju desne spirale sa zajedničkom osom. Jedan zavoj spirale je 10 nukleotida, veličina zavoja je 3,4 nm, visina svakog nukleotida je 0,34 nm, prečnik spirale je 2,0 nm. Kao rezultat rotacije jednog lanca oko drugog, formiraju se glavni žljeb (oko 20 Å u prečniku) i manji žleb (oko 12 Å u prečniku) dvostruke spirale DNK. Ovaj oblik Watson-Crickove dvostruke spirale kasnije je nazvan B-oblik. U ćelijama DNK obično postoji u B obliku, koji je najstabilniji.
Funkcije DNK
Predloženi model objasnio je mnoga biološka svojstva deoksiribonukleinske kiseline, uključujući skladištenje genetskih informacija i raznolikost gena koju obezbjeđuje širok izbor uzastopnih kombinacija od 4 nukleotida i činjenicu postojanja genetskog koda, sposobnost samoreprodukcije. i prenose genetske informacije dobijene procesom replikacije, te implementaciju genetskih informacija u obliku proteina, kao i svih drugih spojeva formiranih uz pomoć proteina enzima.
Osnovne funkcije DNK.
- DNK je nosilac genetske informacije, što je osigurano činjenicom postojanja genetskog koda.
- Reprodukcija i prijenos genetskih informacija kroz generacije ćelija i organizama. Ovu funkcionalnost osigurava proces replikacije.
- Implementacija genetskih informacija u obliku proteina, kao i svih drugih spojeva formiranih uz pomoć proteina enzima. Ovu funkciju pružaju procesi transkripcije i prevođenja.

Oblici organizacije dvolančane DNK
DNK može formirati nekoliko tipova dvostrukih spirala (slika 4). Trenutno je već poznato šest oblika (od A do E i Z-forme).
Strukturni oblici DNK, kako je ustanovila Rosalind Franklin, zavise od zasićenosti molekula nukleinske kiseline vodom. U studijama DNK vlakana uz pomoć rendgenske difrakcijske analize, pokazalo se da rendgenski uzorak radikalno zavisi od relativne vlažnosti pri kojem stepenu zasićenosti vodom ovog vlakna se eksperiment odvija. Ako je vlakno bilo dovoljno zasićeno vodom, tada se dobija jedna radiografija. Kada se osušio, pojavio se potpuno drugačiji rendgenski uzorak, vrlo različit od rendgenskog uzorka vlakana visoke vlage.
Molekul DNK visoke vlažnosti naziva se B-oblik. U fiziološkim uslovima (niska koncentracija soli, visok stepen hidratacije), dominantan strukturni tip DNK je B-oblik (glavni oblik dvolančane DNK - Watson-Crick model). Korak spirale takvog molekula je 3,4 nm. Postoji 10 komplementarnih parova po okretu u obliku upletenih hrpa "kovanica" - azotnih baza. Hrpe se drže zajedno vodoničnim vezama između dva suprotna "kovačića" naslaga, a "namotane" su pomoću dvije trake fosfodiestarske kičme uvijene u desnu spiralu. Ravnine azotnih baza su okomite na osu spirale. Susedni komplementarni parovi su rotirani jedan u odnosu na drugi za 36°. Prečnik spirale je 20Å, sa purinskim nukleotidom 12Å i pirimidinskim 8Å.
Molekul DNK niže vlažnosti naziva se A-oblik. A-oblik nastaje u uslovima manje visoke hidratacije i pri većem sadržaju Na+ ili K+ jona. Ova šira desna spiralna konformacija ima 11 parova baza po okretu. Ravnine azotnih baza imaju jači nagib prema osi heliksa, odstupaju od normale na os heliksa za 20°. To implicira prisustvo unutrašnje praznine prečnika 5Å. Udaljenost između susjednih nukleotida je 0,23 nm, dužina zavoja je 2,5 nm, a promjer spirale je 2,3 nm.
U početku se smatralo da je A oblik DNK manje važan. Međutim, kasnije je postalo jasno da A-forma DNK, kao i B-oblika, ima ogroman biološki značaj. RNA-DNK spirala u kompleksu šablon-prajmer ima A-oblik, kao i RNA-RNA spirala i RNA ukosne strukture (2'-hidroksilna grupa riboze sprečava molekule RNK da formiraju B-oblik). A-oblik DNK nalazi se u sporama. Utvrđeno je da je A-oblik DNK 10 puta otporniji na UV zrake od B-oblika.
A-oblik i B-oblik se nazivaju kanonskim oblicima DNK.
Obrasci C-E takođe dešnjaci, njihovo formiranje se može posmatrati samo u posebnim eksperimentima, i, očigledno, ne postoje in vivo. C oblik DNK ima strukturu sličnu B DNK. Broj parova baza po zavoju je 9,33, dužina zavoja heliksa je 3,1 nm. Parovi baza su nagnuti pod uglom od 8 stepeni u odnosu na okomitu poziciju na osu. Žljebovi su slične veličine žljebovima B-DNK. U ovom slučaju glavni žlijeb je nešto plići, a manji žljeb dublji. Prirodni i sintetički DNK polinukleotidi mogu se transformirati u C-oblik.
| Tabela 1. Karakteristike nekih tipova DNK struktura | |||
| Spiralni tip | A | B | Z |
| Spiral pitch | 0,32 nm | 3,38 nm | 4,46 nm |
| Spiral twist | U redu | U redu | lijevo |
| Broj parova baza po okretu | 11 | 10 | 12 |
| Udaljenost između osnovnih ravnina | 0,256 nm | 0,338 nm | 0,371 nm |
| Konformacija glikozidne veze | anti | anti | anti-C sin-G |
| Konformacija furanoznog prstena | C3"-endo | C2"-endo | C3"-endo-G C2"-endo-C |
| Širina utora, mala/velika | 1,11/0,22 nm | 0,57/1,17 nm | 0,2/0,88 nm |
| Dubina utora, mala/velika | 0,26/1,30 nm | 0,82/0,85 nm | 1,38/0,37 nm |
| Prečnik spirale | 2,3 nm | 2,0 nm | 1,8 nm |
Strukturni elementi DNK
(nekanonske strukture DNK)
Strukturni elementi DNK uključuju neobične strukture ograničene nekim posebnim sekvencama:
|

DNK u obliku slova Z otkriven je 1979. godine tokom proučavanja heksanukleotida d(CG)3 -. Otkrili su ga MIT profesor Alexander Rich i njegove kolege. Z-oblik je postao jedan od najvažnijih strukturnih elemenata DNK zbog činjenice da je njegovo formiranje uočeno u DNK regijama gdje se purini izmjenjuju s pirimidinima (na primjer, 5'-GCGCGC-3'), ili u ponavljanjima 5 '-CGCGCG-3' koji sadrži metilovani citozin. Bitan uslov za formiranje i stabilizaciju Z-DNK bilo je prisustvo purinskih nukleotida u njoj u sin konformaciji, naizmenično sa pirimidinskim bazama u anti konformaciji.
Prirodni molekuli DNK uglavnom postoje u desnorukom B-obliku osim ako ne sadrže sekvence poput (CG)n. Međutim, ako su takve sekvence dio DNK, onda se ovi dijelovi, kada se promijeni ionska snaga otopine ili kationa koji neutraliziraju negativni naboj na fosfodiesterskom okviru, ovi dijelovi mogu transformirati u Z-oblik, dok ostali dijelovi DNK u lanac ostaje u klasičnom B obliku. Mogućnost takvog prijelaza ukazuje na to da su dva lanca u dvostrukoj spirali DNK u dinamičkom stanju i da se mogu odmotati jedan u odnosu na drugi, prelazeći iz desnog oblika u lijevu formu i obrnuto. Biološke posljedice takve labilnosti, koja omogućava konformacijske transformacije strukture DNK, još uvijek nisu u potpunosti shvaćene. Vjeruje se da dijelovi Z-DNK igraju određenu ulogu u regulaciji ekspresije određenih gena i učestvuju u genetskoj rekombinaciji.
Z-oblik DNK je lijeva dvostruka spirala u kojoj je fosfodiesterska kičma smještena u cik-cak uzorku duž osi molekula. Otuda i naziv molekula (cik-cak)-DNK. Z-DNK je najmanje uvrnuta (12 parova baza po okretu) i najtanja DNK poznata u prirodi. Udaljenost između susjednih nukleotida je 0,38 nm, dužina zavoja je 4,56 nm, a prečnik Z-DNK je 1,8 nm. Osim toga, izgled ove molekule DNK odlikuje se prisustvom jednog žlijeba.
Z oblik DNK je pronađen u prokariotskim i eukariotskim ćelijama. Sada su dobijena antitijela koja mogu razlikovati Z-oblik od B-oblika DNK. Ova antitela se vezuju za određene regione ogromnih hromozoma ćelija pljuvačne žlezde Drosophile (Dr. melanogaster). Reakciju vezivanja je lako pratiti zbog neobične strukture ovih hromozoma, u kojoj gušće regije (diskovi) u kontrastu s manje gustim regijama (interdiskovi). Z-DNK regioni se nalaze u interdiskovima. Iz ovoga proizilazi da Z-oblik zapravo postoji u prirodnim uvjetima, iako su veličine pojedinih dijelova Z-oblike još uvijek nepoznate.

(invertori) su najpoznatije i najčešće prisutne bazne sekvence u DNK. Palindrom je riječ ili fraza koja se čita isto s lijeva na desno i obrnuto. Primjeri takvih riječi ili fraza su: KOLIBA, KOZAK, POPLAVA I RUŽA JE PALA NA AZOROVU ŠAPU. Kada se primjenjuje na dijelove DNK, ovaj termin (palindrom) označava istu izmjenu nukleotida duž lanca s desna na lijevo i s lijeva na desno (kao slova u riječi "koliba" itd.).
Palindrom je karakteriziran prisustvom invertiranih ponavljanja baznih sekvenci koje imaju simetriju drugog reda u odnosu na dva lanca DNK. Takve sekvence su, iz očiglednih razloga, samokomplementarne i imaju tendenciju formiranja ukosnih ili krstastih struktura (Sl.). Ukosnice pomažu regulatornim proteinima da prepoznaju gdje je kopiran genetski tekst hromozomske DNK.
Kada je na istom lancu DNK prisutan obrnuti ponavljanje, sekvenca se naziva zrcalni ponavljanje. Ponavljanja ogledala nemaju svojstva samokomplementarnosti i stoga nisu u stanju da formiraju ukosne ili kruciformne strukture. Sekvence ovog tipa nalaze se u gotovo svim velikim DNK molekulima i mogu se kretati od samo nekoliko parova baza do nekoliko hiljada parova baza.
Prisustvo palindroma u obliku kruciformnih struktura u eukariotskim ćelijama nije dokazano, iako je određeni broj kruciformnih struktura otkriven in vivo u ćelijama E. coli. Prisustvo samokomplementarnih sekvenci u RNK ili jednolančanoj DNK glavni je razlog savijanja lanca nukleinske kiseline u rastvorima u određenu prostornu strukturu, koju karakteriše formiranje mnogih „ukosnica“.

H-oblika DNK je spirala formirana od tri lanca DNK - trostruka spirala DNK. To je kompleks Watson-Crickove dvostruke spirale sa trećim jednolančanim DNK lancem, koji se uklapa u njegov glavni žljeb, formirajući takozvani Hoogsteen par.
Formiranje takvog tripleksa nastaje kao rezultat savijanja dvostruke spirale DNK na način da polovina njegovog dijela ostaje u obliku dvostruke spirale, a druga polovina je odvojena. U ovom slučaju, jedna od nepovezanih spirala formira novu strukturu s prvom polovinom dvostruke spirale - trostruku spiralu, a druga se ispostavlja nestrukturirana, u obliku jednolančane sekcije. Karakteristika ovog strukturnog prijelaza je njegova oštra ovisnost o pH medija, čiji protoni stabiliziraju novu strukturu. Zbog ove karakteristike, nova struktura je nazvana H-oblik DNK, čije je formiranje otkriveno u supersmotanim plazmidima koji sadrže homopurin-homopirimidinske regije, koji su zrcalni ponavljanje.
U daljnjim istraživanjima ustanovljeno je da je moguće izvršiti strukturnu tranziciju nekih homopurin-homopirimidin dvolančanih polinukleotida sa formiranjem trolančane strukture koja sadrži:
- jedan homopurinski i dva homopirimidinska lanca ( Py-Pu-Py triplex) [Hoogsteen interakcija].
Sastavni blokovi Py-Pu-Py tripleksa su kanonske izomorfne CGC+ i TAT trijade. Za stabilizaciju tripleksa potrebna je protonacija CGC+ trijade, tako da ovi tripleksi zavise od pH rastvora.
- jedan homopirimidin i dva homopurinska lanca ( Py-Pu-Pu triplex) [inverzna Hoogsteenova interakcija].
Sastavni blokovi Py-Pu-Pu tripleksa su kanonske izomorfne CGG i TAA trijade. Bitno svojstvo Py-Pu-Pu tripleksa je ovisnost njihove stabilnosti o prisustvu dvostruko nabijenih jona, a za stabilizaciju tripleksa različitih sekvenci potrebni su različiti ioni. Budući da formiranje Py-Pu-Pu tripleksa ne zahtijeva protoniranje njihovih sastavnih nukleotida, takvi tripleksi mogu postojati pri neutralnom pH.
Napomena: direktne i reverzne Hoogsteen interakcije se objašnjavaju simetrijom 1-metiltimina: rotacija od 180° dovodi do toga da atom O2 zauzima mjesto atoma O4, dok je sistem vodoničnih veza očuvan.
Poznata su dva tipa trostrukih spirala:
- paralelne trostruke spirale u kojima se polaritet trećeg lanca poklapa sa polaritetom homopurinskog lanca Watson-Crickovog dupleksa
- antiparalelne trostruke spirale, u kojima su polaritet trećeg i homopurinskog lanca suprotni.

G-kvadrupleks- 4-lančani DNK. Ova struktura nastaje ako postoje četiri guanina, koji formiraju takozvani G-kvadrupleks - okrugli ples od četiri guanina.
Prvi nagovještaji mogućnosti formiranja takvih struktura dobili su mnogo prije probojnog rada Watsona i Cricka - davne 1910. godine. Tada je njemački hemičar Ivar Bang otkrio da jedna od komponenti DNK - gvanozinska kiselina - formira gelove u visokim koncentracijama, dok druge komponente DNK nemaju ovo svojstvo.
Godine 1962. metodom difrakcije rendgenskih zraka bilo je moguće utvrditi ćelijsku strukturu ovog gela. Ispostavilo se da se sastoji od četiri ostatka gvanina, koji se međusobno povezuju u krug i formiraju karakterističan kvadrat. U centru vezu podržava jon metala (Na, K, Mg). Iste strukture se mogu formirati u DNK ako sadrži puno gvanina. Ovi ravni kvadrati (G-kvarteti) su naslagani da formiraju prilično stabilne, guste strukture (G-kvadrupleksi).
Četiri odvojena lanca DNK mogu se utkati u četverolančane komplekse, ali ovo je prilično izuzetak. Češće se jedan lanac nukleinske kiseline jednostavno veže u čvor, formirajući karakteristična zadebljanja (na primjer, na krajevima hromozoma), ili dvolančana DNK na nekom području bogatom gvaninom formira lokalni kvadrupleks.
Najviše je proučavano postojanje kvadrupleksa na krajevima hromozoma - na telomerima i u tumorskim promotorima. Međutim, potpuna slika lokalizacije takve DNK u ljudskim hromozomima još uvijek nije poznata.
Sve ove neobične strukture DNK u linearnom obliku su nestabilne u poređenju sa B-formom DNK. Međutim, DNK često postoji u kružnom obliku topološke napetosti kada ima ono što se naziva supersmotanjem. Pod ovim uslovima lako se formiraju nekanonske DNK strukture: Z-oblici, „križevi” i „ukosnice”, H-oblici, gvaninski kvadrupleksi i i-motiv.
- Superzamotani oblik – primećuje se kada se oslobodi iz jezgra ćelije bez oštećenja pentozofosfatne kičme. Ima oblik super-upletenih zatvorenih prstenova. U supernamotanom stanju, dvostruka spirala DNK se barem jednom „uvrće u sebe“, odnosno sadrži najmanje jedan superokret (poprimi oblik osmice).
- Opušteno stanje DNK - posmatrano sa jednim prekidom (prekid jednog lanca). U ovom slučaju, superzavojnice nestaju i DNK poprima oblik zatvorenog prstena.
- Linearni oblik DNK se opaža kada su dva lanca dvostruke spirale prekinuta.
Tercijarna struktura DNK
Tercijarna struktura DNK nastaje kao rezultat dodatnog uvijanja u prostoru dvostruke spiralne molekule - njenog supersmotanja. Supersmotavanje molekula DNK u eukariotskim ćelijama, za razliku od prokariota, javlja se u obliku kompleksa sa proteinima.
Gotovo sav DNK eukariota nalazi se u hromozomima jezgara; samo mala količina sadržana je u mitohondrijima, a u biljkama u plastidima. Glavna tvar hromozoma eukariotskih ćelija (uključujući ljudske hromozome) je kromatin, koji se sastoji od dvolančane DNK, histona i nehistonskih proteina.
Proteini histonskog hromatina
Histoni su jednostavni proteini koji čine do 50% hromatina. U svim proučavanim životinjskim i biljnim ćelijama pronađeno je pet glavnih klasa histona: H1, H2A, H2B, H3, H4, koji se razlikuju po veličini, sastavu aminokiselina i naboju (uvijek pozitivno).
Histon H1 sisara sastoji se od jednog polipeptidnog lanca koji sadrži približno 215 aminokiselina; veličine ostalih histona variraju od 100 do 135 aminokiselina. Svi su spiralizirani i uvijeni u globulu prečnika oko 2,5 nm, a sadrže neobično veliku količinu pozitivno nabijenih aminokiselina lizina i arginina. Histoni mogu biti acetilirani, metilirani, fosforilirani, poli(ADP)-ribozilirani, a histoni H2A i H2B su kovalentno vezani za ubikvitin. Uloga takvih modifikacija u formiranju strukture i obavljanju funkcija histona još nije u potpunosti razjašnjena. Pretpostavlja se da je to njihova sposobnost da komuniciraju sa DNK i obezbede jedan od mehanizama za regulaciju delovanja gena.
Histoni stupaju u interakciju s DNK prvenstveno putem jonskih veza (mostova soli) formiranih između negativno nabijenih fosfatnih grupa DNK i pozitivno nabijenih lizinskih i argininskih ostataka histona.
Nehistonski hromatinski proteini
Nehistonski proteini, za razliku od histona, veoma su raznoliki. Izolovano je do 590 različitih frakcija nehistonskih proteina koji se vezuju za DNK. Nazivaju se i kiselim proteinima, jer njihovom strukturom dominiraju kisele aminokiseline (oni su polianioni). Raznolikost nehistonskih proteina povezana je sa specifičnom regulacijom aktivnosti hromatina. Na primjer, enzimi potrebni za replikaciju i ekspresiju DNK mogu se privremeno vezati za kromatin. Drugi proteini, recimo, oni koji su uključeni u različite regulatorne procese, vezuju se za DNK samo u određenim tkivima ili u određenim fazama diferencijacije. Svaki protein je komplementaran specifičnoj sekvenci DNK nukleotida (DNK mjesto). Ova grupa uključuje:
- porodica proteina cinkovog prsta specifičnih za lokaciju. Svaki „cink prst“ prepoznaje određeno mjesto koje se sastoji od 5 parova nukleotida.
- porodica proteina specifičnih za lokaciju - homodimeri. Fragment takvog proteina u kontaktu s DNK ima strukturu spirala-zavoj-heliks.
- gel proteini visoke pokretljivosti (HMG proteini) su grupa strukturnih i regulatornih proteina koji su stalno povezani sa hromatinom. Imaju molekularnu težinu manju od 30 kDa i karakteriše ih visok sadržaj nabijenih aminokiselina. Zbog svoje male molekularne težine, HMG proteini imaju veliku pokretljivost tokom elektroforeze u poliakrilamidnom gelu.
- enzimi replikacije, transkripcije i popravke.
Uz učešće strukturnih, regulatornih proteina i enzima uključenih u sintezu DNK i RNK, nit nukleosoma se pretvara u visoko kondenzovani kompleks proteina i DNK. Dobivena struktura je 10.000 puta kraća od originalne DNK molekule.

hromatin
Kromatin je kompleks proteina s nuklearnom DNK i neorganskim tvarima. Većina hromatina je neaktivna. Sadrži čvrsto zbijenu, kondenzovanu DNK. Ovo je heterohromatin. Postoje konstitutivni, genetski neaktivni hromatin (satelitska DNK) koji se sastoje od neizraženih regiona, i fakultativni - neaktivni u nizu generacija, ali pod određenim okolnostima sposoban za ekspresiju.
Aktivni hromatin (euhromatin) je nekondenzovan, tj. manje zbijeno. U različitim ćelijama njegov sadržaj se kreće od 2 do 11%. U ćelijama mozga ga ima najviše - 10-11%, u ćelijama jetre - 3-4 i ćelijama bubrega - 2-3%. Uočena je aktivna transkripcija euhromatina. Štaviše, njegova strukturna organizacija omogućava da se iste genetske informacije o DNK svojstvene određenom tipu organizma koriste drugačije u specijalizovanim ćelijama.
U elektronskom mikroskopu, slika hromatina podsjeća na perle: sferna zadebljanja veličine oko 10 nm, odvojena mostovima nalik na niti. Ova sferna zadebljanja nazivaju se nukleosomi. Nukleosom je strukturna jedinica hromatina. Svaki nukleosom sadrži supernamotani DNK segment od 146 bp koji formira 1,75 lijevog zavoja po nukleosomskom jezgru. Nukleosomsko jezgro je histonski oktamer koji se sastoji od histona H2A, H2B, H3 i H4, po dva molekula svakog tipa (slika 9), koji izgleda kao disk prečnika 11 nm i debljine 5,7 nm. Peti histon, H1, nije dio nukleozomskog jezgra i nije uključen u proces namotavanja DNK na histonski oktamer. Kontaktira DNK na mjestima gdje dvostruka spirala ulazi i izlazi iz nukleosomskog jezgra. To su intercore (linker) sekcije DNK, čija dužina varira u zavisnosti od tipa ćelije od 40 do 50 parova nukleotida. Kao rezultat toga, dužina fragmenta DNK uključenog u nukleozome također varira (od 186 do 196 parova nukleotida).
Nukleozomi sadrže otprilike 90% DNK, a ostatak su linkeri. Vjeruje se da su nukleosomi fragmenti "tihog" kromatina, a linker je aktivan. Međutim, nukleosomi se mogu razviti i postati linearni. Nesavijeni nukleosomi su već aktivni kromatin. Ovo jasno pokazuje zavisnost funkcije od strukture. Može se pretpostaviti da što je više hromatina sadržano u globularnim nukleosomima, to je manje aktivan. Očigledno je da je u različitim stanicama nejednak udio kromatina u mirovanju povezan s brojem takvih nukleosoma.
Na elektronskim mikroskopskim fotografijama, u zavisnosti od uslova izolacije i stepena rastezanja, kromatin može izgledati ne samo kao duga nit sa zadebljanjima - „zrncima“ nukleozoma, već i kao kraća i gušća fibrila (vlakna) prečnika od 30 nm, čije se formiranje uočava tokom interakcije histona H1 vezanog za vezni region DNK i histona H3, što dovodi do dodatnog uvrtanja heliksa od šest nukleozoma po okretu kako bi se formirao solenoid prečnika 30 nm. U ovom slučaju, histonski protein može ometati transkripciju brojnih gena i tako regulirati njihovu aktivnost.
Kao rezultat interakcije DNK sa histonima opisanim gore, segment dvostruke spirale DNK od 186 parova baza prosječnog prečnika 2 nm i dužine 57 nm pretvara se u spiralu prečnika 10 nm i dužine 5 nm. Kada se ova spirala naknadno sabije u vlakno prečnika 30 nm, stepen kondenzacije se povećava još šest puta.
Konačno, pakovanje DNK dupleksa sa pet histona rezultira 50-strukom kondenzacijom DNK. Međutim, čak ni tako visok stepen kondenzacije ne može objasniti skoro 50.000 - 100.000 puta zbijanje DNK u metafaznom hromozomu. Nažalost, detalji daljeg pakovanja hromatina do metafaznog hromozoma još nisu poznati, tako da možemo samo razmotriti opšte karakteristike ovog procesa.
Nivoi zbijenosti DNK u hromozomima
Svaki molekul DNK je upakovan u poseban hromozom. Ljudske diploidne ćelije sadrže 46 hromozoma, koji se nalaze u ćelijskom jezgru. Ukupna dužina DNK svih hromozoma u ćeliji je 1,74 m, ali je prečnik jezgra u koji su hromozomi upakovani milionima puta manji. Takvo kompaktno pakovanje DNK u hromozomima i hromozomima u ćelijskom jezgru je obezbeđeno raznim histonskim i nehistonskim proteinima koji u određenoj sekvenci interaguju sa DNK (vidi gore). Zbijanje DNK u hromozomima omogućava smanjenje njenih linearnih dimenzija za približno 10.000 puta - otprilike od 5 cm do 5 mikrona. Postoji nekoliko nivoa zbijenosti (slika 10).


- Dvostruka spirala DNK je negativno nabijena molekula promjera 2 nm i dužine od nekoliko cm.
- nivo nukleozoma- hromatin u elektronskom mikroskopu izgleda kao lanac "perli" - nukleozoma - "na niti". Nukleosom je univerzalna strukturna jedinica koja se nalazi i u euhromatinu i u heterohromatinu, u interfaznom jezgru i metafaznim hromozomima.
Nukleosomski nivo zbijenosti osiguravaju posebni proteini - histoni. Osam pozitivno nabijenih histonskih domena formiraju jezgro nukleosoma oko kojeg je namotana negativno nabijena DNK molekula. To daje skraćenje od 7 puta, dok se prečnik povećava sa 2 na 11 nm.
- nivo solenoida
Solenoidni nivo organizacije hromozoma karakteriše se uvijanjem filamenta nukleosoma i formiranjem debljih fibrila prečnika 20-35 nm - solenoida ili superbida. Korak solenoida je 11 nm; ima oko 6-10 nukleozoma po okretu. Solenoidno pakovanje se smatra verovatnijim od superbid pakovanja, prema kojem je hromatinska fibrila prečnika 20-35 nm lanac granula, odnosno superbida, od kojih se svaki sastoji od osam nukleozoma. Na nivou solenoida, linearna veličina DNK se smanjuje za 6-10 puta, promjer se povećava na 30 nm.
- nivo petlje
Nivo petlje osiguravaju DNK-vezujući proteini koji nisu specifični za histonsko mjesto, koji prepoznaju i vezuju se za specifične sekvence DNK, formirajući petlje od približno 30-300 kb. Petlja osigurava ekspresiju gena, tj. petlja nije samo strukturna, već i funkcionalna formacija. Skraćivanje na ovom nivou se dešava 20-30 puta. Prečnik se povećava na 300 nm. U citološkim preparatima mogu se vidjeti strukture u obliku petlje, poput „četkica lampe“ u oocitima vodozemaca. Čini se da su ove petlje superzamotane i predstavljaju domene DNK, vjerovatno koje odgovaraju jedinicama transkripcije i replikacije hromatina. Specifični proteini fiksiraju baze petlji i, moguće, neke od njihovih unutrašnjih dijelova. Organizacija domena nalik petlji promovira savijanje hromatina u metafaznim hromozomima u spiralne strukture višeg reda.
- nivo domena
Nivo domena organizacije hromozoma nije dovoljno proučavan. Na ovom nivou uočava se formiranje domena petlje - strukture niti (vlakna) debljine 25-30 nm, koje sadrže 60% proteina, 35% DNK i 5% RNK, praktično su nevidljive u svim fazama ćelijskog ciklusa sa izuzetak mitoze i donekle su nasumično raspoređeni po ćelijskom jezgru. U citološkim preparatima mogu se vidjeti strukture u obliku petlje, poput „četkica lampe“ u oocitima vodozemaca.
Domeni petlje su vezani na svojoj bazi za intranuklearni proteinski matriks na takozvanim ugrađenim mjestima vezivanja, koji se često nazivaju MAR/SAR sekvence (MAR, iz engleskog matriks povezanog regiona; SAR, iz engleskog područja vezanja skele) - DNK fragmenti dužine nekoliko stotina parova baza koje karakteriše visok sadržaj (>65%) A/T nukleotidnih parova. Čini se da svaka domena ima jedno porijeklo replikacije i funkcionira kao autonomna superhelikalna jedinica. Bilo koja domena petlje sadrži mnogo transkripcionih jedinica čije je funkcionisanje vjerovatno koordinisano - cijeli domen je ili u aktivnom ili neaktivnom stanju.
Na nivou domena, kao rezultat sekvencijalnog pakovanja hromatina, dolazi do smanjenja linearnih dimenzija DNK za približno 200 puta (700 nm).
- hromozomskom nivou
Na hromozomskom nivou, kondenzacija hromozoma profaze u metafazni hromozom se dešava sa zbijanjem domena petlje oko aksijalnog okvira nehistonskih proteina. Ovo supersmotanje je praćeno fosforilacijom svih H1 molekula u ćeliji. Kao rezultat toga, metafazni hromozom se može prikazati kao gusto zbijene solenoidne petlje, umotane u čvrstu spiralu. Tipični ljudski hromozom može sadržavati do 2.600 petlji. Debljina takve strukture dostiže 1400 nm (dvije hromatide), a molekula DNK se skraćuje 104 puta, tj. od 5 cm rastegnute DNK do 5 µm.
Funkcije hromozoma
U interakciji sa ekstrahromozomskim mehanizmima, hromozomi obezbeđuju
- pohranjivanje nasljednih informacija
- koristeći ove informacije za stvaranje i održavanje ćelijske organizacije
- regulisanje čitanja nasljednih informacija
- samoumnožavanje genetskog materijala
- transfer genetskog materijala sa ćelije majke na ćelije kćeri.
Postoje dokazi da kada se aktivira regija hromatina, tj. tokom transkripcije, prvo histon H1, a zatim histonski oktet se reverzibilno uklanjaju iz njega. To uzrokuje dekondenzaciju hromatina, sekvencijalni prijelaz 30-nm hromatinske fibrile u 10-nm fibril i njeno dalje odvijanje u dijelove slobodne DNK, tj. gubitak strukture nukleozoma.
Nakon otkrića principa molekularne organizacije takve supstance kao što je DNK 1953. godine, počela se razvijati molekularna biologija. Dalje u procesu istraživanja, naučnici su otkrili kako se DNK rekombinuje, njen sastav i kako je strukturiran naš ljudski genom.
Svakodnevno se dešavaju složeni procesi na molekularnom nivou. Kako je strukturiran molekul DNK, od čega se sastoji? I kakvu ulogu imaju molekuli DNK u ćeliji? Razgovarajmo detaljno o svim procesima koji se odvijaju unutar dvostrukog lanca.
Šta je nasledna informacija?
Pa gdje je sve počelo? Davne 1868. pronašli su ga u jezgrima bakterija. A 1928. N. Koltsov je iznio teoriju da je u DNK da su sve genetske informacije o živom organizmu šifrirane. Tada su J. Watson i F. Crick 1953. godine pronašli model danas dobro poznate DNK heliksa, za koji su zasluženo dobili priznanje i nagradu – Nobelovu nagradu.
Šta je uopšte DNK? Ova tvar se sastoji od 2 ujedinjene niti, odnosno spirale. Dio takvog lanca sa određenim informacijama naziva se gen.

DNK pohranjuje sve informacije o tome koje vrste proteina će se formirati i kojim redoslijedom. Makromolekula DNK je materijalni nosilac nevjerovatno obimnih informacija, koje se bilježe u strogom nizu pojedinačnih cigli - nukleotida. Ukupno ima 4 nukleotida koji se međusobno nadopunjuju hemijski i geometrijski. Ovaj princip komplementarnosti, ili komplementarnosti, u nauci će biti opisan kasnije. Ovo pravilo igra ključnu ulogu u kodiranju i dekodiranju genetskih informacija.
Pošto je lanac DNK nevjerovatno dugačak, u ovoj sekvenci nema ponavljanja. Svako živo biće ima svoj jedinstveni lanac DNK.
Funkcije DNK
Funkcije uključuju skladištenje nasljednih informacija i njihovo prenošenje na potomstvo. Bez ove funkcije, genom vrste se ne bi mogao očuvati i razvijati hiljadama godina. Organizmi koji su prošli teške mutacije gena vjerojatnije neće preživjeti ili izgubiti sposobnost da proizvode potomstvo. Tako nastaje prirodna zaštita od degeneracije vrste.

Još jedna bitna funkcija je implementacija pohranjenih informacija. Ćelija ne može stvoriti niti jedan vitalni protein bez tih instrukcija koje su pohranjene u dvostrukom lancu.
Sastav nukleinske kiseline
Sada se pouzdano zna od čega se sastoje sami nukleotidi - građevni blokovi DNK. Sadrže 3 supstance:
- Ortofosforna kiselina.
- Azotna baza. Pirimidinske baze - koje imaju samo jedan prsten. To uključuje timin i citozin. Purinske baze, koje sadrže 2 prstena. To su gvanin i adenin.
- Saharoza. DNK sadrži deoksiribozu, RNK ribozu.
Broj nukleotida je uvijek jednak broju azotnih baza. U posebnim laboratorijama nukleotid se razgrađuje i iz njega se izoluje dušična baza. Tako se proučavaju pojedinačna svojstva ovih nukleotida i moguće mutacije u njima.
Nivoi organizacije nasljednih informacija
Postoje 3 nivoa organizacije: genetski, hromozomski i genomski. Sve informacije potrebne za sintezu novog proteina sadržane su u malom dijelu lanca - genu. Odnosno, gen se smatra najnižim i najjednostavnijim nivoom kodiranja informacija.

Geni se pak sklapaju u hromozome. Zahvaljujući ovakvoj organizaciji nosioca nasljednog materijala, grupe osobina se izmjenjuju po određenim zakonima i prenose s jedne generacije na drugu. Treba napomenuti da u tijelu postoji nevjerovatan broj gena, ali se informacija ne gubi ni kada se rekombinuje mnogo puta.
Postoji nekoliko vrsta gena:
- Prema funkcionalnoj namjeni, razlikuju se 2 tipa: strukturne i regulatorne sekvence;
- Na osnovu njihovog uticaja na procese koji se odvijaju u ćeliji razlikuju se: supervitalni, smrtonosni, uslovno smrtonosni geni, kao i mutatorski i antimutatorski geni.
Geni su raspoređeni duž hromozoma u linearnom redu. U hromozomima informacije nisu nasumično fokusirane, postoji određeni redosled. Postoji čak i mapa koja pokazuje položaje ili lokuse gena. Na primjer, poznato je da hromozom broj 18 šifrira podatke o boji djetetovih očiju.
Šta je genom? Ovo je naziv dat čitavom nizu nukleotidnih sekvenci u ćeliji organizma. Genom karakterizira cijelu vrstu, a ne pojedinca.
Šta je ljudski genetski kod?
Činjenica je da sav ogroman potencijal ljudskog razvoja leži već u periodu začeća. Sve nasljedne informacije koje su neophodne za razvoj zigota i rast djeteta nakon rođenja šifrirane su u genima. DNK dijelovi su najosnovniji nosioci nasljednih informacija.
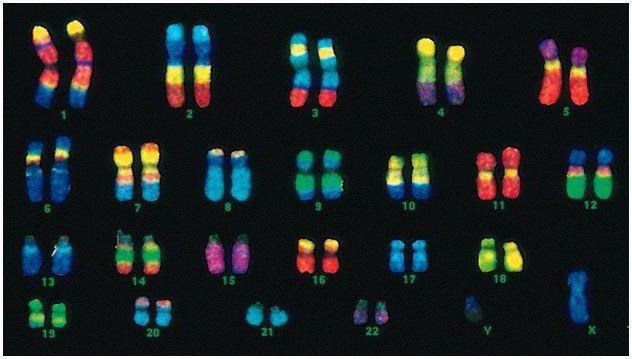
Ljudi imaju 46 hromozoma, ili 22 somatska para plus jedan hromozom koji određuje pol od svakog roditelja. Ovaj diploidni skup hromozoma kodira cjelokupni fizički izgled čovjeka, njegove mentalne i fizičke sposobnosti i podložnost bolestima. Somatski hromozomi se spolja ne razlikuju, ali nose različite informacije, jer je jedan od oca, drugi od majke.
Muški kod se razlikuje od ženskog po zadnjem paru hromozoma - XY. Ženski diploidni set je posljednji par, XX. Muškarci dobijaju jedan X hromozom od svoje biološke majke, koji se potom prenosi na njihove kćeri. Spolni Y hromozom se prenosi na sinove.
Ljudski hromozomi se jako razlikuju po veličini. Na primjer, najmanji par hromozoma je broj 17. A najveći par je 1 i 3.
Promjer dvostruke spirale kod ljudi je samo 2 nm. DNK je tako čvrsto smotana da se uklapa u malo jezgro ćelije, iako bi bila duga i do 2 metra ako bi se odmotala. Dužina spirale je stotine miliona nukleotida.
Kako se genetski kod prenosi?
Dakle, kakvu ulogu imaju molekuli DNK u diobi ćelija? Geni - nosioci nasljednih informacija - nalaze se unutar svake ćelije tijela. Kako bi prenijeli svoj kod na organizam kćeri, mnoga stvorenja dijele svoju DNK na 2 identična spirala. Ovo se zove replikacija. Tokom procesa replikacije, DNK se odmotava i posebne "mašine" dovršavaju svaki lanac. Nakon što se genetski heliks račva, jezgro i sve organele počinju da se dijele, a zatim i cijela stanica.
Ali ljudi imaju drugačiji proces prenošenja gena – seksualni. Karakteristike oca i majke su pomiješane, novi genetski kod sadrži podatke oba roditelja.
Čuvanje i prijenos nasljednih informacija je moguć zbog složene organizacije spirale DNK. Uostalom, kao što smo rekli, struktura proteina je šifrirana u genima. Jednom stvoren u vrijeme začeća, ovaj kod će se kopirati kroz život. Kariotip (lični skup hromozoma) se ne menja tokom obnavljanja ćelija organa. Prijenos informacija vrši se uz pomoć polnih spolnih stanica - muških i ženskih.
Samo virusi koji sadrže jedan lanac RNK nisu sposobni prenijeti svoje informacije svom potomstvu. Stoga su im potrebne ljudske ili životinjske stanice za reprodukciju.
Implementacija nasljednih informacija
U ćelijskom jezgru se stalno odvijaju važni procesi. Sve informacije zabilježene u hromozomima koriste se za izgradnju proteina iz aminokiselina. Ali lanac DNK nikada ne napušta jezgro, pa mu je potrebna pomoć još jednog važnog spoja: RNK. RNK je ta koja je u stanju da prodre kroz nuklearnu membranu i stupi u interakciju sa lancem DNK.
Kroz interakciju DNK i 3 vrste RNK, sve kodirane informacije se realizuju. Na kom nivou se dešava implementacija naslednih informacija? Sve interakcije se dešavaju na nivou nukleotida. Messenger RNA kopira dio lanca DNK i dovodi ovu kopiju do ribosoma. Ovdje počinje sinteza nove molekule iz nukleotida.
Da bi mRNA kopirala potrebni dio lanca, spirala se odvija, a zatim se, po završetku procesa rekodiranja, ponovo obnavlja. Štaviše, ovaj proces se može odvijati istovremeno na 2 strane 1 hromozoma.
Princip komplementarnosti
Sastoje se od 4 nukleotida - adenina (A), gvanina (G), citozina (C), timina (T). Oni su povezani vodoničnim vezama prema pravilu komplementarnosti. Rad E. Chargaffa pomogao je da se uspostavi ovo pravilo, budući da je naučnik uočio neke obrasce u ponašanju ovih supstanci. E. Chargaff je otkrio da je molarni omjer adenina i timina jednak jedan. I na isti način, omjer gvanina i citozina uvijek je jednak jedan.
Na osnovu njegovog rada, genetičari su formirali pravilo za interakciju nukleotida. Pravilo komplementarnosti kaže da se adenin kombinuje samo sa timinom, a gvanin samo sa citozinom. Tokom dekodiranja heliksa i sinteze novog proteina u ribosomu, ovo pravilo alternacije pomaže da se brzo pronađe potrebna aminokiselina koja je vezana za prijenosnu RNK.
RNK i njeni tipovi
Šta je nasledna informacija? nukleotidi u dvostrukom lancu DNK. Šta je RNK? Šta je njen posao? RNK, ili ribonukleinska kiselina, pomaže u izdvajanju informacija iz DNK, dekodiranju i, na osnovu principa komplementarnosti, stvaranju proteina neophodnih za ćelije.
Ukupno postoje 3 tipa RNK. Svaki od njih obavlja striktno svoju funkciju.
- Informativni (mRNA), ili se još naziva i matrica. Ide pravo u centar ćelije, u jezgro. Pronalazi u jednom od hromozoma neophodan genetski materijal za izgradnju proteina i kopira jednu od strana dvostrukog lanca. Kopiranje se ponovo dešava po principu komplementarnosti.
- Transport je mali molekul koji na jednoj strani ima dekodere nukleotida, a na drugoj aminokiseline koje odgovaraju osnovnom kodu. Zadatak tRNA je da je dostavi u "radionicu", odnosno u ribozom, gdje sintetiše potrebnu aminokiselinu.
- rRNA je ribosomska. On kontrolira količinu proteina koja se proizvodi. Sastoji se od 2 dijela - aminokiselinskog i peptidnog dijela.
Jedina razlika u dekodiranju je u tome što RNK nema timin. Umjesto timina, ovdje je prisutan uracil. Ali tada, tokom procesa sinteze proteina, tRNA i dalje ispravno instalira sve aminokiseline. Ako dođe do bilo kakvih grešaka u dekodiranju informacija, tada dolazi do mutacije.
Popravak oštećene DNK molekule
Proces obnavljanja oštećenog dvostrukog lanca naziva se popravak. Tokom procesa popravke, oštećeni geni se uklanjaju.

Zatim se traženi redoslijed elemenata tačno reproducira i reže na isto mjesto na lancu odakle je uklonjen. Sve se to događa zahvaljujući posebnim hemikalijama - enzimima.
Zašto dolazi do mutacija?
Zašto neki geni počinju mutirati i prestaju obavljati svoju funkciju – pohranjivanje vitalnih nasljednih informacija? Ovo se događa zbog greške u dekodiranju. Na primjer, ako se adenin slučajno zamijeni timinom.
Postoje i hromozomske i genomske mutacije. Kromosomske mutacije nastaju kada se dijelovi nasljednih informacija izgube, dupliciraju ili čak prenesu i umetnu u drugi kromosom.

Genomske mutacije su najozbiljnije. Njihov uzrok je promjena u broju hromozoma. Odnosno, kada je umjesto para - diploidnog skupa u kariotipu prisutan triploidni skup.
Najpoznatiji primjer triploidne mutacije je Downov sindrom, kod kojeg je lični skup hromozoma 47. Kod takve djece se formiraju 3 hromozoma umjesto 21. para.
Poznata je i mutacija koja se zove poliploidija. Ali poliploidija se javlja samo u biljkama.
Molekul DNK se sastoji od dva lanca koji formiraju dvostruku spiralu. Njegovu strukturu prvi su dešifrovali Francis Crick i James Watson 1953. godine.
U početku je molekul DNK, koji se sastojao od para nukleotidnih lanaca upletenih jedan oko drugog, doveo do pitanja zašto je imao ovakav oblik. Naučnici ovu pojavu nazivaju komplementarnošću, što znači da se samo određeni nukleotidi mogu naći jedan naspram drugog u njegovim lancima. Na primjer, adenin je uvijek suprotan timinu, a gvanin je uvijek suprotan citozinu. Ovi nukleotidi molekule DNK nazivaju se komplementarni.
Šematski je to prikazano ovako:
T - A
C - G
Ovi parovi formiraju hemijsku nukleotidnu vezu, koja određuje redosled aminokiselina. U prvom slučaju je malo slabiji. Veza između C i G je jača. Nekomplementarni nukleotidi ne formiraju parove jedni s drugima.
O zgradi
Dakle, struktura molekula DNK je posebna. Ovakav oblik ima s razlogom: činjenica je da je broj nukleotida vrlo velik i da je potrebno puno prostora za smještaj dugih lanaca. Iz tog razloga lanci se odlikuju spiralnim uvijanjem. Ovaj fenomen se naziva spiralizacija, omogućava da se niti skrate za oko pet do šest puta.
Tijelo koristi neke molekule ove vrste vrlo aktivno, druge rijetko. Potonji, osim spiralizacije, prolaze i kroz takvo "kompaktno pakovanje" kao što je superspiralizacija. I tada se dužina molekule DNK smanjuje za 25-30 puta.

Šta je "pakovanje" molekula?
Proces supersmotanja uključuje histonske proteine. Imaju strukturu i izgled kalema konca ili šipke. Na njih su namotane spiralne niti, koje se odmah "kompaktno upakuju" i zauzimaju malo prostora. Kada se pojavi potreba za korištenjem jedne ili druge niti, ona se odmotava od koluta, na primjer, histonskog proteina, a spirala se odmotava u dva paralelna lanca. Kada je molekul DNK u ovom stanju, iz njega se mogu očitati potrebni genetski podaci. Međutim, postoji jedan uslov. Dobivanje informacija moguće je samo ako struktura molekule DNK ima neuvijeni oblik. Kromosomi koji su dostupni za čitanje nazivaju se euhromatini, a ako su superzamotani, onda su već heterohromatini.
Nukleinske kiseline
Nukleinske kiseline, kao i proteini, su biopolimeri. Glavna funkcija je skladištenje, implementacija i prijenos nasljednih (genetskih informacija). Dolaze u dvije vrste: DNK i RNA (deoksiribonukleinska i ribonukleinska). Monomeri u njima su nukleotidi, od kojih svaki sadrži ostatak fosforne kiseline, petougljični šećer (deoksiriboza/riboza) i dušičnu bazu. DNK kod uključuje 4 vrste nukleotida - adenin (A) / gvanin (G) / citozin (C) / timin (T). Razlikuju se po dušičnoj bazi koju sadrže.
U molekulu DNK broj nukleotida može biti ogroman - od nekoliko hiljada do desetina i stotina miliona. Takvi džinovski molekuli mogu se ispitati putem elektronskog mikroskopa. U ovom slučaju, moći ćete vidjeti dvostruki lanac polinukleotidnih lanaca, koji su međusobno povezani vodoničnim vezama dušičnih baza nukleotida.

Istraživanja
Tokom istraživanja, naučnici su otkrili da se tipovi molekula DNK razlikuju u različitim živim organizmima. Također je utvrđeno da se gvanin jednog lanca može vezati samo za citozin, a timin za adenin. Raspored nukleotida u jednom lancu striktno odgovara paralelnom. Zahvaljujući ovoj komplementarnosti polinukleotida, molekula DNK je sposobna za udvostručenje i samoreprodukciju. Ali prvo se komplementarni lanci, pod utjecajem posebnih enzima koji uništavaju uparene nukleotide, razilaze, a zatim u svakom od njih počinje sinteza lanca koji nedostaje. To se događa zbog slobodnih nukleotida prisutnih u velikim količinama u svakoj ćeliji. Kao rezultat toga, umjesto "majčinog molekula", formiraju se dvije "kćerke", identične po sastavu i strukturi, a DNK kod postaje izvorni. Ovaj proces je preteča ćelijske diobe. Osigurava prijenos svih nasljednih podataka sa matičnih ćelija na ćelije kćeri, kao i na sve naredne generacije.
Kako se čita genski kod?
Danas se ne izračunava samo masa molekula DNK - moguće je saznati i složenije podatke koji su ranije bili nedostupni naučnicima. Na primjer, možete pročitati informacije o tome kako organizam koristi vlastitu ćeliju. Naravno, u početku su ove informacije u kodiranom obliku i imaju oblik određene matrice, te se stoga moraju prenijeti na poseban nosač, a to je RNK. Ribonukleinska kiselina je u stanju da prodre u ćeliju kroz nuklearnu membranu i pročita kodirane informacije iznutra. Dakle, RNK je nosilac skrivenih podataka od jezgra do ćelije, a razlikuje se od DNK po tome što umesto dezoksiriboze sadrži ribozu i uracil umesto timina. Osim toga, RNK je jednolančana.

RNA sinteza
Dubinska analiza DNK pokazala je da nakon što RNK napusti jezgro, ona ulazi u citoplazmu, gdje se može integrirati kao matriks u ribozome (posebne enzimske sisteme). Vođeni primljenim informacijama, oni mogu sintetizirati odgovarajući slijed proteinskih aminokiselina. Ribosom uči iz trojnog koda koji tip organskog jedinjenja treba da bude vezan za formirajući proteinski lanac. Svaka aminokiselina ima svoj specifični triplet, koji je kodira.
Nakon što je formiranje lanca završeno, on dobija specifičan prostorni oblik i pretvara se u protein sposoban da obavlja svoje hormonalne, građevinske, enzimske i druge funkcije. Za svaki organizam to je genski proizvod. Iz njega se određuju sve vrste kvaliteta, svojstava i manifestacija gena.

Geni
Procesi sekvenciranja prvenstveno su razvijeni da bi se dobile informacije o tome koliko gena ima molekul DNK u svojoj strukturi. I, iako su istraživanja omogućila naučnicima da naprave veliki napredak po ovom pitanju, još nije moguće znati njihov tačan broj.
Prije samo nekoliko godina pretpostavljalo se da molekuli DNK sadrže otprilike 100 hiljada gena. Nešto kasnije, brojka se smanjila na 80 hiljada, a 1998. godine genetičari su izjavili da je u jednoj DNK prisutno samo 50 hiljada gena, što je samo 3% ukupne dužine DNK. Ali najnoviji zaključci genetičara bili su upečatljivi. Sada tvrde da genom uključuje 25-40 hiljada ovih jedinica. Ispostavilo se da je samo 1,5% hromozomske DNK odgovorno za kodiranje proteina.
Istraživanje se tu nije zaustavilo. Paralelni tim stručnjaka za genetski inženjering otkrio je da je broj gena u jednom molekulu tačno 32 hiljade. Kao što vidite, još uvijek je nemoguće dobiti konačan odgovor. Previše je kontradikcija. Svi istraživači se oslanjaju samo na svoje rezultate.
Da li je postojala evolucija?
Unatoč činjenici da nema dokaza o evoluciji molekula (pošto je struktura molekule DNK krhka i male veličine), naučnici su ipak napravili jednu pretpostavku. Na osnovu laboratorijskih podataka, iznijeli su sljedeću verziju: u početnoj fazi svog pojavljivanja, molekula je imala oblik jednostavnog samoreplicirajućeg peptida, koji je uključivao do 32 aminokiseline pronađene u drevnim oceanima.
Nakon samoreplikacije, zahvaljujući silama prirodne selekcije, molekuli su stekli sposobnost da se zaštite od vanjskih elemenata. Počele su da žive duže i da se razmnožavaju u većim količinama. Molekuli koji su se našli u lipidnom mehuru imali su sve šanse da se sami reprodukuju. Kao rezultat niza uzastopnih ciklusa, lipidni mjehurići su dobili oblik ćelijskih membrana, a zatim - dobro poznatih čestica. Treba napomenuti da je danas bilo koji dio molekule DNK složena i jasno funkcionalna struktura, čije sve karakteristike naučnici još nisu u potpunosti proučili.

Moderni svijet
Nedavno su naučnici iz Izraela razvili kompjuter koji može da izvrši trilione operacija u sekundi. Danas je to najbrži automobil na Zemlji. Cijela tajna je da inovativni uređaj pokreće DNK. Profesori kažu da će u bliskoj budućnosti takvi računari čak moći da generišu energiju.
Prije godinu dana stručnjaci sa Weizman instituta u Rehovotu (Izrael) najavili su stvaranje programabilne molekularne računarske mašine koja se sastoji od molekula i enzima. S njima su zamijenili silikonske mikročipove. Do danas, tim je dodatno napredovao. Sada samo jedan molekul DNK može da obezbedi kompjuteru potrebne podatke i potrebno gorivo.
Biohemijski „nanokompjuteri” nisu fikcija, već postoje u prirodi i manifestuju se u svakom živom biću. Ali često njima ne upravljaju ljudi. Osoba još ne može da operiše genom bilo koje biljke kako bi izračunala, recimo, broj „Pi“.
Na ideju o korištenju DNK za pohranjivanje/obradu podataka naučnici su prvi put pali na pamet 1994. godine. Tada je molekul korišten za rješavanje jednostavnog matematičkog problema. Od tada, brojne istraživačke grupe su predložile različite projekte vezane za DNK kompjutere. Ali ovdje su se svi pokušaji zasnivali samo na energetskom molekulu. Takav kompjuter ne možete vidjeti golim okom, on izgleda kao prozirna otopina vode u epruveti. U njemu nema mehaničkih dijelova, već samo trilioni biomolekularnih uređaja - i to samo u jednoj kapi tečnosti!
Ljudski DNK
Ljudi su postali svjesni vrste ljudske DNK 1953. godine, kada su naučnici prvi put mogli svijetu demonstrirati model dvolančane DNK. Za to su Kirk i Votson dobili Nobelovu nagradu, jer je ovo otkriće postalo fundamentalno u 20. veku.
Vremenom su, naravno, dokazali da strukturirani ljudski molekul može izgledati ne samo kao u predloženoj verziji. Nakon detaljnije analize DNK, otkrili su A-, B- i ljevoruki oblik Z-. Forma A- često je izuzetak, jer nastaje samo ako postoji nedostatak vlage. Ali to je moguće samo u laboratorijskim studijama; za prirodno okruženje to je anomalno; takav proces se ne može dogoditi u živoj ćeliji.
B-oblik je klasičan i poznat je kao dvostruki desnoruki lanac, ali Z-oblik nije samo uvrnut u suprotnom smjeru ulijevo, već ima i više cik-cak izgled. Naučnici su takođe identifikovali oblik G-kvadrupleksa. Njegova struktura nema 2, već 4 niti. Prema genetičarima, ovaj oblik se javlja u područjima gdje postoji višak gvanina.

Veštački DNK
Danas već postoji veštačka DNK, koja je identična kopija pravog; savršeno prati strukturu prirodne dvostruke spirale. Ali, za razliku od originalnog polinukleotida, umjetni ima samo dva dodatna nukleotida.
Pošto je sinkronizacija napravljena na osnovu informacija dobijenih iz različitih studija stvarne DNK, ona se takođe može kopirati, samoreplicirati i razvijati. Stručnjaci su radili na stvaranju takvog umjetnog molekula oko 20 godina. Rezultat je nevjerovatan izum koji može koristiti genetski kod na isti način kao prirodni DNK.
Četiri postojeće azotne baze genetičari su dodali još dvije, koje su nastale hemijskom modifikacijom prirodnih baza. Za razliku od prirodne DNK, ispostavilo se da je umjetna DNK prilično kratka. Sadrži samo 81 bazni par. Međutim, on se također razmnožava i razvija.
Replikacija molekula dobijenog umjetnim putem odvija se zahvaljujući lančanoj reakciji polimeraze, ali do sada se to ne događa samostalno, već uz intervenciju naučnika. Oni samostalno dodaju potrebne enzime navedenoj DNK, stavljajući je u posebno pripremljenu tečnu podlogu.
Konačan rezultat
Na proces i konačni ishod razvoja DNK mogu uticati različiti faktori, kao što su mutacije. Zbog toga je neophodno proučavati uzorke materije kako bi rezultat analize bio pouzdan i pouzdan. Primjer je test očinstva. Ali ne možemo a da se ne radujemo što su incidenti poput mutacije rijetki. Ipak, uzorci materije se uvijek iznova provjeravaju kako bi se na osnovu analize dobile tačnije informacije.

Biljni DNK
Zahvaljujući visokim tehnologijama sekvenciranja (HTS), napravljena je revolucija u oblasti genomike – moguća je i ekstrakcija DNK iz biljaka. Naravno, dobijanje visokokvalitetne DNK molekulske mase iz biljnog materijala predstavlja određene poteškoće zbog velikog broja kopija DNK mitohondrija i hloroplasta, kao i visokog nivoa polisaharida i fenolnih jedinjenja. Za izolaciju strukture koju razmatramo u ovom slučaju, koriste se različite metode.
Vodikova veza u DNK
Vodikova veza u molekuli DNK odgovorna je za elektromagnetsko privlačenje stvoreno između pozitivno nabijenog atoma vodika koji je vezan za elektronegativni atom. Ova dipolna interakcija ne ispunjava kriterijum hemijske veze. Ali može se pojaviti intermolekularno ili u različitim dijelovima molekule, tj. intramolekularno.

Atom vodika vezuje se za elektronegativni atom koji je donor veze. Elektronegativni atom može biti dušik, fluor ili kisik. Ona – kroz decentralizaciju – privlači oblak elektrona iz jezgre vodika na sebe i čini atom vodika (djelimično) pozitivno nabijenim. Budući da je veličina H-a mala u poređenju s drugim molekulima i atomima, naboj je također mali.
DNK dekodiranje
Pre nego što dešifruju molekul DNK, naučnici prvo uzmu ogroman broj ćelija. Za što precizniji i uspješniji rad potrebno ih je oko milion. Rezultati dobijeni tokom istraživanja se stalno upoređuju i bilježe. Danas dekodiranje genoma više nije rijetkost, već pristupačna procedura.

Naravno, dešifrovanje genoma jedne ćelije je nepraktična vežba. Podaci dobijeni tokom ovakvih studija nisu od interesa za naučnike. Ali važno je shvatiti da sve trenutno postojeće metode dekodiranja, uprkos njihovoj složenosti, nisu dovoljno efikasne. Oni će omogućiti čitanje samo 40-70% DNK.
Međutim, profesori sa Harvarda nedavno su najavili metodu pomoću koje se može dešifrirati 90% genoma. Tehnika se zasniva na dodavanju molekula prajmera u izolovane ćelije, uz pomoć kojih počinje replikacija DNK. Ali čak se i ova metoda ne može smatrati uspješnom; još uvijek je treba usavršiti prije nego što se može otvoreno koristiti u nauci.
