The DNA molecule is located. DNA and genes. New look at DNA
Biochemical bases of heredity.
Genetic role of nucleic acids.
Nucleic acids are biological polymers found in all cells, from primitive to complex ones. They were first discovered by Johann Friedrich Miescher in 1868 in cells rich in nuclear material (leukocytes, salmon sperm). The term “nucleic acids” was proposed in 1889.
There are two types of nucleic acids: DNA, RNA (ATP - mononucleotide). DNA and RNA are molecules - matrices. DNA is contained about 6 * 10 -12 g in somatic cells: in the nucleus, mitochondria. RNA is part of ribosomes and is found in the nucleus and cytoplasm.
The study and proof of the leading role of nucleic acids in the transmission of hereditary information was carried out on viral particles. Tobacco mosaic virus is known to be virulent for tobacco and plantain. The viral particle consists of 95% protein and 5% nucleic acid. The protein capsid in the viral particles was swapped, but after some time the protein in both strains was transformed into its previous form.
In bacteriophages that infect E. coli, the phage coat proteins were labeled with radioactive S, and the phage DNA was labeled with radioactive P. In a bacterial cell infected with the phage, phage particles were formed that contained only radioactive P.
Structure and functions of DNA and RNA molecules.
Nucleic acids are biopolymers of irregular structure, the monomers of which are nucleotides. The nucleotide consists of residues of three substances: phosphoric acid, carbohydrate - pentose, nitrogenous base. DNA nucleotides contain the carbohydrate deoxyribose, and RNA contains ribose. Residues of purine and pyrimidine nitrogenous bases that make up DNA are adenine, guanine, cytosine, thymine. RNA molecules contain adenine, guanine, cytosine, and uracil.
Nucleotides are connected to each other through the phosphoric acid residue of one nucleotide and the carbohydrate of another by a strong covalent ester bond, called an “oxygen bridge.” The bond goes through the 5th carbon atom of a carbohydrate of one nucleotide to the 3rd carbon atom of a carbohydrate of another nucleotide. The nucleotide sequence represents the primary structure of nucleic acids. RNA is a single polynucleotide chain. DNA's structure is a double polynucleotide chain, coiled into a spiral.
The secondary structure of DNA is formed when a second DNA strand appears, arranged according to the principle of complementarity with respect to the first. The second chain is opposite to the first (antiparallel). The nitrogenous bases lie in a plane perpendicular to the plane of the molecule - this resembles a spiral staircase. The railings of this staircase are phosphoric acid residues and carbohydrates, and the steps are nitrogenous bases.
The nitrogenous bases that make up each nucleotide in oppositely directed chains are capable of forming complementary hydrogen bonds with each other (due to the existing functional groups in the structure of each nitrogenous base). Adenyl nucleotide is complementary to thymine, guanyl to cytosine, and vice versa. By themselves, these bonds are fragile, but a DNA molecule “stitched” repeatedly along its entire length with such bonds represents a very strong connection.
Complementarity- this is the spatial-structural and chemical correspondence of nitrogenous bases to each other; they fit each other “like a key to a lock.”
One DNA molecule can contain 10 8 or more nucleotides.
The structure of the DNA molecule as a double antiparallel helix was proposed in 1953 by the American biologist James Watson and the English physicist Francis Crick.
The DNA molecule of any living organism on the planet consists of only four types of nucleotides, differing from each other in the nitrogenous bases they contain: adenyl, guanyl, thymine and cytosine. In that versatility DNA. Their sequence is different, and the number is infinite.
For each species of living organisms and for each organism separately, this sequence is individual and strictly specific .
Peculiarity The structure of DNA is that the chemically active sections of the molecule - nitrogenous bases, are immersed in the center of the helix and form complementary bonds with each other, and deoxyribose and phosphoric acid residues are located on the periphery and cover access to nitrogenous bases - they are chemically inactive. This structure can maintain chemical stability for a long time. What else is needed to store hereditary information? It is these features of DNA structure that determine its ability to encode and reproduce genetic information.
The strong structure of DNA is quite difficult to destroy. Nevertheless, this happens regularly in the cell - during the synthesis of RNA and the doubling of the DNA molecule itself before cell division.
Duplication, DNA replication begins with the fact that a special enzyme - DNA polymerase - unwinds the double helix and separates it into separate strands - a reduplication fork is formed. The enzyme acts like a lock on a zipper. At each single-stranded chain - the sticky ends of the reduplication fork - a new chain is synthesized from free nucleotides in the karyoplasm according to the principle of complementarity. In two new DNA molecules, one strand remains the original mother strand, and the second remains the new daughter strand. As a result, instead of one DNA molecule, two molecules appear with the same exact nucleotide composition as the original one.
In living systems we encounter a new type of reactions unknown in inanimate nature. They're called matrix synthesis reactions . Matrix synthesis is like casting on a matrix: new molecules are synthesized in exact accordance with the plan laid down in the structure of existing molecules. These reactions ensure the exact sequence of monomer units in the synthesized polymers. Monomers arrive at a specific location on molecules that serve as a matrix where the reaction takes place. If such reactions occurred as a result of random collisions of molecules, they would proceed infinitely slowly. The synthesis of complex molecules based on the template principle is carried out quickly and accurately with the help of enzymes. Template synthesis underlies the most important reactions in the synthesis of nucleic acids and proteins. The role of the matrix in the cell is played by molecules of nucleic acids DNA or RNA. Monomeric molecules from which the polymer is synthesized - nucleotides or amino acids - in accordance with the principle of complementarity, are located and fixed on the matrix in a strictly defined order. Then the monomer units are connected into a polymer chain, and the finished polymer leaves the matrix. After this, the matrix is ready for the assembly of a new, exactly the same polymer molecule.
Matrix-type reactions are a specific feature of a living cell. They are the basis of the fundamental property of all living things - the ability to reproduce their own kind.
Functions of nucleic acids– storage and transmission of hereditary information. DNA molecules encode information about the primary structure of the protein. The synthesis of mRNA molecules takes place on the DNA matrix. This process is called "transcription". I-RNA in the process of “translation” implements information in the form of a sequence of amino acids in a protein molecule.
The DNA of each cell carries information not only about the structural proteins that determine the shape of the cell, but also about all enzyme proteins, hormone proteins and other proteins, as well as the structure of all types of RNA.
Perhaps nucleic acids provide various types of biological memory - immunological, neurological, etc., and also play a significant role in the regulation of biosynthetic processes.
Related information.
The first proof of the role of DNA as a carrier of hereditary information in organisms attracted enormous attention to the study of nucleic acids. In 1869, F. Miescher isolated a special substance from cell nuclei, which he called nuclein. After 20 years this name was replaced by the term nucleic acid. In 1924, R. Felgen developed a method for the cytological recognition of nucleic acids through their specific staining and showed that DNA is localized in the nuclei of cells, and RNA in the cytoplasm. In 1936 A.N. Belozersky and I.I. Dubrovskaya isolated DNA in its pure form from the nuclei of plant cells. By the beginning of the 1930s. The basic chemical principles of the structure of nucleic acid sugars were elucidated, and in 1953 a structural model of DNA was created.
The basic structural unit of nucleic acids is nucleotide, which consists of three chemically different parts connected by covalent bonds (Fig. 5.2).
Rice. 5.2. Structural formulas: A- nucleotides; b- DNA; V - RNA (see also p. 110)

Rice. 5.2. Ending. Structural formulas: A- nucleotides; 6 - DNA; V- RNA
The first part is sugar containing five carbon atoms: deoxyribose in DNA and ribose in RNA.
The second part of the nucleotide, a purine or pyrimidine nitrogenous base, is covalently linked to the first carbon atom of the sugar, forming a structure called nucleoside. DNA contains purine bases - adenine(A) and guanine(D) - and pyrimidine bases - thymine(T) and cytosine(C). The corresponding nucleosides are called deoxyadenosine, deoxyguanosine, deoxythymidine and deoxycytidine. RNA contains the same purine bases as DNA, a pyrimidine base cytosine, and instead of thymine it contains uracil(U); the corresponding nucleosides are called adenosine, guanosine, uridine and cytidine.
The third part of the nucleotide is a phosphate group, which connects neighboring nucleosides into a polymer chain through phosphodiester bonds between the 5-carbon atom of one sugar and the 3-carbon atom of another (Fig. 5.2, b, V). Nucleotides are called nucleosides with one or more phosphate groups attached by ester bonds to the 3" or 5 carbon atoms of the sugar. The synthesis of nucleotides precedes the synthesis of nucleic acids; accordingly, nucleotides are products of chemical or enzymatic hydrolysis of nucleic acids.
Nucleic acids are very long polymer chains consisting of mononucleotides connected by 5- and 3'-phosphodiester bonds. An intact DNA molecule contains, depending on the type of organism, from several thousand to many millions of nucleotides, an intact RNA molecule - from 100 to 100 thousand or more nucleotides.
The results of E. Chargaff's analyzes of the nucleotide composition of DNA of various species forms showed that the molecular ratio of various nitrogenous bases - adenine, guanine, thymine, cytosine - varies widely. Consequently, it was proven that DNA is not at all a monotonous polymer consisting of identical tetranucleotides, as was assumed in the 40s. XX centuries, and that it fully possesses the complexity necessary for the preservation and transmission of hereditary information in the form of a specific sequence of nucleotide bases.
Research by E. Chargaff also revealed a feature inherent in all DNA molecules: the molar content of adenine is equal to the content of thymine, and the molar content of guanine is equal to the content of cytosine. These equalities are called Chargaff's equivalence rule: [A] = [T], [G] = [C]; the number of purines is equal to the number of pyrimidines. Depending on the species, only the ratio ([A] + [T])/([G] + [C]) changes (Table 5.1).
|
Composition of bases |
Attitude |
Asymmetry |
|||||
|
reasons |
|||||||
|
(A + T)/(G + C) |
|||||||
|
Animals |
|||||||
|
Turtle |
|||||||
|
sea crab |
|||||||
|
Sea urchin |
|||||||
|
Plants, mushrooms |
|||||||
|
wheat germ |
|||||||
|
Mushroom Aspergillus niger |
|||||||
|
Bacteria |
|||||||
|
Escherichia coli |
|||||||
|
Staphylococcus aureus |
|||||||
|
Clostridium perfringens |
|||||||
|
Brucela abortus |
|||||||
|
Sarcina lutea |
|||||||
|
Bacteriophages |
|||||||
|
FH 174 (viral form) |
|||||||
|
FH 174 (replicative form) |
|||||||
The ratio of bases is called nucleotide coefficient(species) specificity. Chargaff's discovery formulated an important structural feature of DNA, which was later reflected in the structural model of DNA by J. Watson and F. Crick (1953), who actually showed that Chargaff's rules do not impose any restrictions on the possible number of combinations of different base sequences capable of forming molecules DNA.
The concept of nucleotide specificity formed the basis of a new branch of biology - gene systematics, which operates by comparing the composition and structure of nucleic acids to build a natural system of organisms.
According to the Watson-Crick model, a DNA molecule consists of two polynucleotide chains (strands, strands), connected to each other using transverse hydrogen bonds between nitrogenous bases according to the complementary principle (adenine of one chain is connected by two hydrogen bonds with thymine of the opposite chain, and guanine and cytosine of different chains are connected to each other by three hydrogen bonds). In this case, two polynucleotide chains of one molecule are anti-parallel, i.e., opposite the 3" end of one chain is the 5" end of the other chain and vice versa (Fig. 5.3). However, one should keep in mind modern data that the genetic material of some viruses is represented by single-stranded (single-stranded) DNA molecules. Based on X-ray diffraction analysis of DNA, J. Watson and F. Crick also concluded that its double-stranded molecule has a secondary structure in the form of a helix twisted in the direction from left to right, which was later called the 5-form (Fig. 5.4). To date, it has been proven that in addition to the most common 5-form, it is possible to detect DNA sections that have a different configuration - as right-handed (forms A, C), and twisted from right to left (left-twisted, or Z-shape) (Fig. 5.4). There are certain differences between these forms of secondary DNA structure (Table 5.2). For example, the distance between two adjacent pairs of nitrogenous bases in a double-stranded helix, expressed in nanometers (nm), is characterized by different values for the 5-form and Z-form (0.34 and 0.38 nm, respectively). In Fig. Figure 5.5 shows modern three-dimensional models of “left-handed” and “right-handed” forms of DNA.

Rice. 5.3. schematic representation of the primary structure of a fragment of a double-stranded DNA molecule: A - adenine; G - guanine; T - thymine; C - cytosine

Rice. 5.4.

Table 5.2
Properties of various forms of DNA double helices
RNA molecules, depending on their structural and functional characteristics, are divided into several types: messenger RNA (mRNA, or mRNA), ribosomal RNA (rRNA), transfer RNA (tRNA), small nuclear RNA (snRNA), etc. In contrast From DNA, RNA molecules are always single-stranded (single-stranded). However, they can form more complex (secondary) configurations due to the complementary connection of individual sections of such a chain based on the interaction of complementary nitrogenous bases (A-U and G-C). As an example, consider the cloverleaf configuration for the phenylalanine transfer RNA molecule (Figure 5.6).

Rice. 5.6.
In 1953, D. Watson and F. Crick proposed a model of the structure of DNA, which was based on the following postulates:
- 1. DNA is a polymer consisting of nucleotides connected by 3" and 5" phosphodiester bonds.
- 2. The composition of DNA nucleotides obeys Chargaff's rules.
- 3. The DNA molecule has a double helix structure, reminiscent of a spiral staircase, as evidenced by x-ray diffraction patterns of DNA strands first obtained by M. Wilkins and R. Franklin.
- 4. The structure of the polymer, as shown by acid-base titration of native (natural) DNA, is stabilized by hydrogen bonds. Titration and heating of native DNA causes a noticeable change in its physical properties, in particular viscosity, transforming it into a denatured form, and covalent bonds are not destroyed.
Nucleic acids are high-molecular substances consisting of mononucleotides, which are connected to each other in a polymer chain using 3", 5" phosphodiester bonds and are packaged in cells in a certain way.
Nucleic acids are biopolymers of two types: ribonucleic acid (RNA) and deoxyribonucleic acid (DNA). Each biopolymer consists of nucleotides that differ in the carbohydrate residue (ribose, deoxyribose) and one of the nitrogenous bases (uracil, thymine). According to these differences, nucleic acids received their name.
Structure of deoxyribonucleic acid
Nucleic acids have a primary, secondary and tertiary structure.
Primary structure of DNA
The primary structure of DNA is a linear polynucleotide chain in which mononucleotides are connected by 3", 5" phosphodiester bonds. The starting material for the assembly of a nucleic acid chain in a cell is the 5"-triphosphate nucleoside, which, as a result of the removal of β and γ phosphoric acid residues, is capable of attaching the 3" carbon atom of another nucleoside. Thus, the 3" carbon atom of one deoxyribose is covalently linked to the 5" carbon atom of another deoxyribose through a single phosphoric acid residue and forms a linear polynucleotide chain of nucleic acid. Hence the name: 3", 5" phosphodiester bonds. Nitrogen bases do not take part in connecting nucleotides of one chain (Fig. 1.).
Such a connection, between the phosphoric acid molecule residue of one nucleotide and the carbohydrate of another, leads to the formation of a pentose-phosphate skeleton of the polynucleotide molecule, on which nitrogenous bases are attached to the side one after another. Their sequence of arrangement in the chains of nucleic acid molecules is strictly specific for the cells of different organisms, i.e. has a specific character (Chargaff's rule).
A linear DNA chain, the length of which depends on the number of nucleotides included in the chain, has two ends: one is called the 3" end and contains a free hydroxyl, and the other is called the 5" end and contains a phosphoric acid residue. The circuit is polar and can have a direction of 5"->3" and 3"->5". The exception is circular DNA.
The genetic "text" of DNA is composed of code "words" - triplets of nucleotides called codons. Sections of DNA containing information about the primary structure of all types of RNA are called structural genes.
Polynucleotide DNA chains reach gigantic sizes, so they are packaged in a certain way in the cell.
While studying the composition of DNA, Chargaff (1949) established important patterns regarding the content of individual DNA bases. They helped reveal the secondary structure of DNA. These patterns are called Chargaff's rules. Chargaff rules
These rules indicate that when constructing DNA, a fairly strict correspondence (pairing) must be observed not of purine and pyrimidine bases in general, but specifically of thymine with adenine and cytosine with guanine. Based on these rules, in 1953, Watson and Crick proposed a model of the secondary structure of DNA, called the double helix (Fig.). |

Secondary structure of DNA
The secondary structure of DNA is a double helix, the model of which was proposed by D. Watson and F. Crick in 1953.
Prerequisites for creating a DNA model
As a result of initial analyzes, it was believed that DNA of any origin contains all four nucleotides in equal molar quantities. However, in the 1940s, E. Chargaff and his colleagues, as a result of analyzing DNA isolated from a variety of organisms, clearly showed that they contained nitrogenous bases in different quantitative ratios. Chargaff found that although these ratios are the same for DNA from all cells of the same species of organism, DNA from different species can differ markedly in the content of certain nucleotides. This suggested that the differences in the ratio of nitrogenous bases may be associated with some kind of biological code. Although the ratio of individual purine and pyrimidine bases in different DNA samples turned out to be different, when comparing the test results, a certain pattern emerged: in all samples, the total number of purines was equal to the total number of pyrimidines (A + G = T + C), the amount of adenine was equal to the amount of thymine (A = T), and the amount of guanine is the amount of cytosine (G = C). DNA isolated from mammalian cells was generally richer in adenine and thymine and relatively poorer in guanine and cytosine, whereas DNA from bacteria was richer in guanine and cytosine and relatively poorer in adenine and thymine. These data formed an important part of the factual material on the basis of which the Watson-Crick model of DNA structure was later built.
Another important indirect indication of the possible structure of DNA was provided by L. Pauling’s data on the structure of protein molecules. Pauling showed that several different stable configurations of the amino acid chain in a protein molecule are possible. One common peptide chain configuration, the α-helix, is a regular helical structure. With this structure, the formation of hydrogen bonds between amino acids located on adjacent turns of the chain is possible. Pauling described the α-helical configuration of the polypeptide chain in 1950 and suggested that DNA molecules probably have a helical structure held in place by hydrogen bonds.
However, the most valuable information about the structure of the DNA molecule was provided by the results of X-ray diffraction analysis. X-rays passing through a DNA crystal undergo diffraction, that is, they are deflected in certain directions. The degree and nature of the deflection of the rays depend on the structure of the molecules themselves. An X-ray diffraction pattern (Fig. 3) gives the experienced eye a number of indirect indications regarding the structure of the molecules of the substance under study. Analysis of X-ray diffraction patterns of DNA led to the conclusion that the nitrogenous bases (which have a flat shape) are arranged like a stack of plates. X-ray diffraction patterns revealed three main periods in the structure of crystalline DNA: 0.34, 2 and 3.4 nm.

Watson-Crick DNA model
Based on Chargaff's analytical data, Wilkins' X-ray patterns, and the research of chemists who provided information about the precise distances between atoms in a molecule, the angles between the bonds of a given atom, and the size of the atoms, Watson and Crick began to build physical models of the individual components of the DNA molecule at a certain scale and “adjust” them to each other in such a way that the resulting system corresponds to various experimental data [show] .
It was known even earlier that neighboring nucleotides in a DNA chain are connected by phosphodiester bridges, linking the 5"-carbon deoxyribose atom of one nucleotide with the 3"-carbon deoxyribose atom of the next nucleotide. Watson and Crick had no doubt that the period of 0.34 nm corresponds to the distance between successive nucleotides in the DNA chain. Further, it could be assumed that the period of 2 nm corresponds to the thickness of the chain. And in order to explain what real structure the period of 3.4 nm corresponds to, Watson and Crick, as well as Pauling earlier, suggested that the chain is twisted in the form of a spiral (or, more precisely, forms a helical line, since a spiral in the strict sense of this words are obtained when the coils form a conical rather than cylindrical surface in space). Then a period of 3.4 nm will correspond to the distance between successive turns of this helix. Such a spiral can be very dense or somewhat stretched, that is, its turns can be flat or steep. Since the period of 3.4 nm is exactly 10 times the distance between successive nucleotides (0.34 nm), it is clear that each complete turn of the helix contains 10 nucleotides. From these data, Watson and Crick were able to calculate the density of a polynucleotide chain twisted into a helix with a diameter of 2 nm, with a distance between turns of 3.4 nm. It turned out that such a chain would have a density that was half that of the actual density of DNA, which was already known. I had to assume that the DNA molecule consists of two chains - that it is a double helix of nucleotides.
The next task was, of course, to clarify the spatial relationships between the two chains forming the double helix. Having tried a number of options for the arrangement of chains on their physical model, Watson and Crick found that all the available data was best matched by the option in which two polynucleotide helices go in opposite directions; in this case, chains consisting of sugar and phosphate residues form the surface of the double helix, and purines and pyrimidines are located inside. The bases located opposite each other, belonging to two chains, are connected in pairs by hydrogen bonds; It is these hydrogen bonds that hold the chains together, thus fixing the overall configuration of the molecule.
The double helix of DNA can be imagined as a rope ladder that is twisted in a helical manner, so that its rungs remain horizontal. Then the two longitudinal ropes will correspond to chains of sugar and phosphate residues, and the crossbars will correspond to pairs of nitrogenous bases connected by hydrogen bonds.
As a result of further study of possible models, Watson and Crick concluded that each "crossbar" should consist of one purine and one pyrimidine; at a period of 2 nm (corresponding to the diameter of the double helix), there would not be enough space for two purines, and the two pyrimidines could not be close enough to each other to form proper hydrogen bonds. An in-depth study of the detailed model showed that adenine and cytosine, while forming a combination of a suitable size, could still not be positioned in such a way that hydrogen bonds would form between them. Similar reports forced the exclusion of the combination guanine - thymine, while the combinations adenine - thymine and guanine - cytosine turned out to be quite acceptable. The nature of hydrogen bonds is such that adenine forms a pair with thymine, and guanine with cytosine. This idea of specific base pairing made it possible to explain the “Chargaff rule”, according to which in any DNA molecule the amount of adenine is always equal to the content of thymine, and the amount of guanine is always equal to the amount of cytosine. Two hydrogen bonds are formed between adenine and thymine, and three between guanine and cytosine. Due to this specificity, the formation of hydrogen bonds against each adenine in one chain causes thymine to form on the other; in the same way, only cytosine can be opposite each guanine. Thus, the chains are complementary to each other, that is, the sequence of nucleotides in one chain uniquely determines their sequence in the other. The two chains run in opposite directions and their terminal phosphate groups are at opposite ends of the double helix.
As a result of their research, in 1953 Watson and Crick proposed a model of the structure of the DNA molecule (Fig. 3), which remains relevant to the present day. According to the model, the DNA molecule consists of two complementary polynucleotide chains. Each DNA strand is a polynucleotide consisting of several tens of thousands of nucleotides. In it, neighboring nucleotides form a regular pentose-phosphate backbone due to the connection of a phosphoric acid residue and deoxyribose by a strong covalent bond. The nitrogenous bases of one polynucleotide chain are arranged in a strictly defined order opposite the nitrogenous bases of the other. The alternation of nitrogenous bases in a polynucleotide chain is irregular.
The arrangement of nitrogenous bases in the DNA chain is complementary (from the Greek “complement” - addition), i.e. Thymine (T) is always against adenine (A), and only cytosine (C) is against guanine (G). This is explained by the fact that A and T, as well as G and C, strictly correspond to each other, i.e. complement each other. This correspondence is determined by the chemical structure of the bases, which allows the formation of hydrogen bonds in the purine and pyrimidine pair. There are two connections between A and T, and three between G and C. These bonds provide partial stabilization of the DNA molecule in space. The stability of the double helix is directly proportional to the number of G≡C bonds, which are more stable compared to A=T bonds.
The known sequence of arrangement of nucleotides in one DNA chain makes it possible, according to the principle of complementarity, to establish the nucleotides of another chain.
In addition, it has been established that nitrogenous bases having an aromatic structure in an aqueous solution are located one above the other, forming, as it were, a stack of coins. This process of forming stacks of organic molecules is called stacking. The polynucleotide chains of the DNA molecule of the Watson-Crick model under consideration have a similar physicochemical state, their nitrogenous bases are arranged in the form of a stack of coins, between the planes of which van der Waals interactions (stacking interactions) arise.
Hydrogen bonds between complementary bases (horizontally) and stacking interactions between planes of bases in a polynucleotide chain due to van der Waals forces (vertically) provide the DNA molecule with additional stabilization in space.
The sugar phosphate backbones of both chains face outward, and the bases face inward, towards each other. The direction of the chains in DNA is antiparallel (one of them has a direction of 5"->3", the other - 3"->5", i.e. the 3" end of one chain is located opposite the 5" end of the other.). The chains form right-handed spirals with a common axis. One turn of the helix is 10 nucleotides, the size of the turn is 3.4 nm, the height of each nucleotide is 0.34 nm, the diameter of the helix is 2.0 nm. As a result of the rotation of one strand around another, a major groove (about 20 Å in diameter) and a minor groove (about 12 Å in diameter) of the DNA double helix are formed. This form of the Watson-Crick double helix was later called the B-form. In cells, DNA usually exists in the B form, which is the most stable.
Functions of DNA
The proposed model explained many biological properties of deoxyribonucleic acid, including the storage of genetic information and the diversity of genes provided by a wide variety of sequential combinations of 4 nucleotides and the fact of the existence of a genetic code, the ability to self-reproduce and transmit genetic information provided by the replication process, and the implementation of genetic information in the form of proteins, as well as any other compounds formed with the help of enzyme proteins.
Basic functions of DNA.
- DNA is the carrier of genetic information, which is ensured by the fact of the existence of a genetic code.
- Reproduction and transmission of genetic information across generations of cells and organisms. This functionality is provided by the replication process.
- Implementation of genetic information in the form of proteins, as well as any other compounds formed with the help of enzyme proteins. This function is provided by the processes of transcription and translation.

Forms of organization of double-stranded DNA
DNA can form several types of double helices (Fig. 4). Currently, six forms are already known (from A to E and Z-form).
The structural forms of DNA, as Rosalind Franklin established, depend on the saturation of the nucleic acid molecule with water. In studies of DNA fibers using X-ray diffraction analysis, it was shown that the X-ray pattern radically depends on the relative humidity at what degree of water saturation of this fiber the experiment takes place. If the fiber was sufficiently saturated with water, then one radiograph was obtained. When dried, a completely different X-ray pattern appeared, very different from the X-ray pattern of high-moisture fiber.
High humidity DNA molecule is called B-form. Under physiological conditions (low salt concentration, high degree of hydration), the dominant structural type of DNA is the B-form (the main form of double-stranded DNA - the Watson-Crick model). The helix pitch of such a molecule is 3.4 nm. There are 10 complementary pairs per turn in the form of twisted stacks of “coins” - nitrogenous bases. The stacks are held together by hydrogen bonds between two opposing “coins” of the stacks, and are “wound” by two ribbons of phosphodiester backbone twisted into a right-handed helix. The planes of the nitrogenous bases are perpendicular to the axis of the helix. Adjacent complementary pairs are rotated relative to each other by 36°. The diameter of the helix is 20Å, with the purine nucleotide occupying 12Å and the pyrimidine nucleotide 8Å.
The lower humidity DNA molecule is called A-form. The A-form is formed under conditions of less high hydration and at a higher content of Na + or K + ions. This broader right-handed helical conformation has 11 base pairs per turn. The planes of the nitrogenous bases have a stronger inclination to the helix axis; they are deviated from the normal to the helix axis by 20°. This implies the presence of an internal void with a diameter of 5Å. The distance between adjacent nucleotides is 0.23 nm, the length of the turn is 2.5 nm, and the diameter of the helix is 2.3 nm.
The A form of DNA was initially thought to be less important. However, it later became clear that the A-form of DNA, like the B-form, has enormous biological significance. The RNA-DNA helix in the template-primer complex has the A-form, as well as the RNA-RNA helix and RNA hairpin structures (the 2'-hydroxyl group of ribose prevents RNA molecules from forming the B-form). The A-form of DNA is found in spores. It has been established that the A-form of DNA is 10 times more resistant to UV rays than the B-form.
The A-form and B-form are called the canonical forms of DNA.
Forms C-E also right-handed, their formation can only be observed in special experiments, and, apparently, they do not exist in vivo. The C form of DNA has a structure similar to B DNA. The number of base pairs per turn is 9.33, the length of the helix turn is 3.1 nm. The base pairs are inclined at an angle of 8 degrees relative to the perpendicular position to the axis. The grooves are similar in size to the grooves of B-DNA. In this case, the main groove is somewhat shallower, and the minor groove is deeper. Natural and synthetic DNA polynucleotides can transform into the C-form.
| Table 1. Characteristics of some types of DNA structures | |||
| Spiral type | A | B | Z |
| Spiral pitch | 0.32 nm | 3.38 nm | 4.46 nm |
| Spiral twist | Right | Right | Left |
| Number of base pairs per turn | 11 | 10 | 12 |
| Distance between base planes | 0.256 nm | 0.338 nm | 0.371 nm |
| Glycosidic bond conformation | anti | anti | anti-C sin-G |
| Conformation of the furanose ring | C3"-endo | C2"-endo | C3"-endo-G C2"-endo-C |
| Groove width, small/large | 1.11/0.22 nm | 0.57/1.17 nm | 0.2/0.88 nm |
| Groove depth, small/large | 0.26/1.30 nm | 0.82/0.85 nm | 1.38/0.37 nm |
| Spiral diameter | 2.3 nm | 2.0 nm | 1.8 nm |
Structural elements of DNA
(non-canonical DNA structures)
The structural elements of DNA include unusual structures limited by some special sequences:
|

Z-shape DNA was discovered in 1979 while studying the hexanucleotide d(CG)3 -. It was discovered by MIT professor Alexander Rich and his colleagues. The Z-form has become one of the most important structural elements of DNA due to the fact that its formation has been observed in DNA regions where purines alternate with pyrimidines (for example, 5'-GCGCGC-3'), or in repeats 5'-CGCGCG-3' containing methylated cytosine. An essential condition for the formation and stabilization of Z-DNA was the presence of purine nucleotides in it in the syn conformation, alternating with pyrimidine bases in the anti conformation.
Natural DNA molecules mainly exist in the right-handed B-form unless they contain sequences like (CG)n. However, if such sequences are part of DNA, then these sections, when the ionic strength of the solution or cations that neutralize the negative charge on the phosphodiester framework changes, these sections can transform into the Z-form, while other sections of DNA in the chain remain in the classical B-form. The possibility of such a transition indicates that the two strands in the DNA double helix are in a dynamic state and can unwind relative to each other, moving from the right-handed form to the left-handed one and vice versa. The biological consequences of such lability, which allows conformational transformations of the DNA structure, are not yet fully understood. It is believed that sections of Z-DNA play a certain role in regulating the expression of certain genes and take part in genetic recombination.
The Z-form of DNA is a left-handed double helix in which the phosphodiester backbone is located in a zigzag pattern along the axis of the molecule. Hence the name of the molecule (zigzag)-DNK. Z-DNA is the least twisted (12 base pairs per turn) and thinnest DNA known in nature. The distance between adjacent nucleotides is 0.38 nm, the length of the turn is 4.56 nm, and the diameter of Z-DNA is 1.8 nm. In addition, the appearance of this DNA molecule is distinguished by the presence of a single groove.
The Z form of DNA has been found in prokaryotic and eukaryotic cells. Antibodies have now been obtained that can distinguish the Z-form from the B-form of DNA. These antibodies bind to certain regions of the giant chromosomes of the salivary gland cells of Drosophila (Dr. melanogaster). The binding reaction is easy to monitor due to the unusual structure of these chromosomes, in which denser regions (disks) contrast with less dense regions (interdisks). Z-DNA regions are located in the interdisks. It follows from this that the Z-form actually exists in natural conditions, although the sizes of individual sections of the Z-form are still unknown.

(inverters) are the most famous and frequently occurring base sequences in DNA. A palindrome is a word or phrase that reads the same from left to right and vice versa. Examples of such words or phrases are: HUT, COSSACK, FLOOD, AND THE ROSE FALLED ON AZOR'S PAW. When applied to DNA sections, this term (palindrome) means the same alternation of nucleotides along the chain from right to left and left to right (like the letters in the word “hut”, etc.).
A palindrome is characterized by the presence of inverted repeats of base sequences that have second-order symmetry relative to two DNA strands. Such sequences, for obvious reasons, are self-complementary and tend to form hairpin or cruciform structures (Fig.). Hairpins help regulatory proteins recognize where the genetic text of chromosome DNA is copied.
When an inverted repeat is present on the same DNA strand, the sequence is called a mirror repeat. Mirror repeats do not have self-complementarity properties and, therefore, are not capable of forming hairpin or cruciform structures. Sequences of this type are found in almost all large DNA molecules and can range from just a few base pairs to several thousand base pairs.
The presence of palindromes in the form of cruciform structures in eukaryotic cells has not been proven, although a certain number of cruciform structures have been detected in vivo in E. coli cells. The presence of self-complementary sequences in RNA or single-stranded DNA is the main reason for the folding of the nucleic acid chain in solutions into a certain spatial structure, characterized by the formation of many “hairpins”.

H-form DNA is a helix formed by three DNA strands - a DNA triple helix. It is a complex of a Watson-Crick double helix with a third single-stranded DNA strand, which fits into its major groove, forming a so-called Hoogsteen pair.
The formation of such a triplex occurs as a result of the folding of a DNA double helix in such a way that half of its section remains in the form of a double helix, and the other half is separated. In this case, one of the disconnected helices forms a new structure with the first half of the double helix - a triple helix, and the second turns out to be unstructured, in the form of a single-stranded section. A feature of this structural transition is its sharp dependence on the pH of the medium, the protons of which stabilize the new structure. Due to this feature, the new structure was called the H-form of DNA, the formation of which was discovered in supercoiled plasmids containing homopurine-homopyrimidine regions, which are a mirror repeat.
In further studies, it was established that it is possible to carry out a structural transition of some homopurine-homopyrimidine double-stranded polynucleotides with the formation of a three-stranded structure containing:
- one homopurine and two homopyrimidine strands ( Py-Pu-Py triplex) [Hoogsteen interaction].
The constituent blocks of the Py-Pu-Py triplex are canonical isomorphic CGC+ and TAT triads. Stabilization of the triplex requires protonation of the CGC+ triad, so these triplexes depend on the pH of the solution.
- one homopyrimidine and two homopurine strands ( Py-Pu-Pu triplex) [inverse Hoogsteen interaction].
The constituent blocks of the Py-Pu-Pu triplex are canonical isomorphic CGG and TAA triads. An essential property of Py-Pu-Pu triplexes is the dependence of their stability on the presence of doubly charged ions, and different ions are required to stabilize triplexes of different sequences. Since the formation of Py-Pu-Pu triplexes does not require protonation of their constituent nucleotides, such triplexes can exist at neutral pH.
Note: direct and reverse Hoogsteen interactions are explained by the symmetry of 1-methylthymine: a rotation of 180° results in the O2 atom taking the place of the O4 atom, while the system of hydrogen bonds is preserved.
Two types of triple helices are known:
- parallel triple helices in which the polarity of the third strand coincides with the polarity of the homopurine chain of the Watson-Crick duplex
- antiparallel triple helices, in which the polarities of the third and homopurine chains are opposite.

G-quadruplex- 4-strand DNA. This structure is formed if there are four guanines, which form the so-called G-quadruplex - a round dance of four guanines.
The first hints of the possibility of the formation of such structures were received long before the breakthrough work of Watson and Crick - back in 1910. Then the German chemist Ivar Bang discovered that one of the components of DNA - guanosinic acid - forms gels at high concentrations, while other components of DNA do not have this property.
In 1962, using the X-ray diffraction method, it was possible to establish the cell structure of this gel. It turned out to be composed of four guanine residues, connecting each other in a circle and forming a characteristic square. In the center, the bond is supported by a metal ion (Na, K, Mg). The same structures can form in DNA if it contains a lot of guanine. These flat squares (G-quartets) are stacked to form fairly stable, dense structures (G-quadruplexes).
Four separate strands of DNA can be woven into four-stranded complexes, but this is rather an exception. More often, a single strand of nucleic acid is simply tied into a knot, forming characteristic thickenings (for example, at the ends of chromosomes), or double-stranded DNA at some guanine-rich region forms a local quadruplex.
The existence of quadruplexes at the ends of chromosomes - at telomeres and in tumor promoters - has been most studied. However, a complete picture of the localization of such DNA in human chromosomes is still not known.
All of these unusual DNA structures in linear form are unstable compared to B-form DNA. However, DNA often exists in a circular form of topological tension when it has what is called supercoiling. Under these conditions, non-canonical DNA structures are easily formed: Z-forms, “crosses” and “hairpins”, H-forms, guanine quadruplexes and i-motif.
- Supercoiled form - noted when released from the cell nucleus without damaging the pentose phosphate backbone. It has the shape of super-twisted closed rings. In the supercoiled state, the DNA double helix is “twisted onto itself” at least once, that is, it contains at least one superturn (takes the shape of a figure eight).
- Relaxed state of DNA - observed with a single break (break of one strand). In this case, the supercoils disappear and the DNA takes the form of a closed ring.
- The linear form of DNA is observed when two strands of a double helix are broken.
Tertiary structure of DNA
Tertiary structure of DNA is formed as a result of additional twisting in space of a double-helical molecule - its supercoiling. Supercoiling of the DNA molecule in eukaryotic cells, unlike prokaryotes, occurs in the form of complexes with proteins.
Almost all of the DNA of eukaryotes is found in the chromosomes of the nuclei; only a small amount is contained in mitochondria, and in plants, in plastids. The main substance of the chromosomes of eukaryotic cells (including human chromosomes) is chromatin, consisting of double-stranded DNA, histone and non-histone proteins.
Histone chromatin proteins
Histones are simple proteins that make up up to 50% of chromatin. In all studied animal and plant cells, five main classes of histones were found: H1, H2A, H2B, H3, H4, differing in size, amino acid composition and charge (always positive).
Mammalian histone H1 consists of a single polypeptide chain containing approximately 215 amino acids; the sizes of other histones vary from 100 to 135 amino acids. All of them are spiralized and twisted into a globule with a diameter of about 2.5 nm, and contain an unusually large amount of positively charged amino acids lysine and arginine. Histones can be acetylated, methylated, phosphorylated, poly(ADP)-ribosylated, and histones H2A and H2B are covalently linked to ubiquitin. The role of such modifications in the formation of the structure and performance of functions by histones has not yet been fully elucidated. It is assumed that this is their ability to interact with DNA and provide one of the mechanisms for regulating gene action.
Histones interact with DNA primarily through ionic bonds (salt bridges) formed between the negatively charged phosphate groups of DNA and the positively charged lysine and arginine residues of histones.
Non-histone chromatin proteins
Non-histone proteins, unlike histones, are very diverse. Up to 590 different fractions of DNA-binding non-histone proteins have been isolated. They are also called acidic proteins, since their structure is dominated by acidic amino acids (they are polyanions). The diversity of non-histone proteins is associated with specific regulation of chromatin activity. For example, enzymes required for DNA replication and expression may bind to chromatin transiently. Other proteins, say, those involved in various regulatory processes, bind to DNA only in specific tissues or at certain stages of differentiation. Each protein is complementary to a specific sequence of DNA nucleotides (DNA site). This group includes:
- family of site-specific zinc finger proteins. Each “zinc finger” recognizes a specific site consisting of 5 nucleotide pairs.
- family of site-specific proteins - homodimers. The fragment of such a protein in contact with DNA has a helix-turn-helix structure.
- high mobility gel proteins (HMG proteins) are a group of structural and regulatory proteins that are constantly associated with chromatin. They have a molecular weight of less than 30 kDa and are characterized by a high content of charged amino acids. Due to their low molecular weight, HMG proteins have high mobility during polyacrylamide gel electrophoresis.
- replication, transcription and repair enzymes.
With the participation of structural, regulatory proteins and enzymes involved in the synthesis of DNA and RNA, the nucleosome thread is converted into a highly condensed complex of proteins and DNA. The resulting structure is 10,000 times shorter than the original DNA molecule.

Chromatin
Chromatin is a complex of proteins with nuclear DNA and inorganic substances. The bulk of the chromatin is inactive. It contains tightly packed, condensed DNA. This is heterochromatin. There are constitutive, genetically inactive chromatin (satellite DNA) consisting of non-expressed regions, and facultative - inactive in a number of generations, but under certain circumstances capable of expression.
Active chromatin (euchromatin) is uncondensed, i.e. packed less tightly. In different cells its content ranges from 2 to 11%. In brain cells it is most abundant - 10-11%, in liver cells - 3-4 and kidney cells - 2-3%. Active transcription of euchromatin is noted. Moreover, its structural organization allows the same genetic DNA information inherent in a given type of organism to be used differently in specialized cells.
In an electron microscope, the image of chromatin resembles beads: spherical thickenings about 10 nm in size, separated by thread-like bridges. These spherical thickenings are called nucleosomes. The nucleosome is a structural unit of chromatin. Each nucleosome contains a 146-bp supercoiled DNA segment wound to form 1.75 left turns per nucleosomal core. The nucleosomal core is a histone octamer consisting of histones H2A, H2B, H3 and H4, two molecules of each type (Fig. 9), which looks like a disk with a diameter of 11 nm and a thickness of 5.7 nm. The fifth histone, H1, is not part of the nucleosomal core and is not involved in the process of winding DNA onto the histone octamer. It contacts DNA at the sites where the double helix enters and exits the nucleosomal core. These are intercore (linker) DNA sections, the length of which varies depending on the cell type from 40 to 50 nucleotide pairs. As a result, the length of the DNA fragment included in the nucleosomes also varies (from 186 to 196 nucleotide pairs).
Nucleosomes contain approximately 90% DNA, the rest being linkers. It is believed that nucleosomes are fragments of “silent” chromatin, and the linker is active. However, nucleosomes can unfold and become linear. Unfolded nucleosomes are already active chromatin. This clearly demonstrates the dependence of function on structure. It can be assumed that the more chromatin is contained in globular nucleosomes, the less active it is. Obviously, in different cells the unequal proportion of resting chromatin is associated with the number of such nucleosomes.
In electron microscopic photographs, depending on the conditions of isolation and the degree of stretching, chromatin can look not only as a long thread with thickenings - “beads” of nucleosomes, but also as a shorter and denser fibril (fiber) with a diameter of 30 nm, the formation of which is observed during interaction histone H1 bound to the linker region of DNA and histone H3, which leads to additional twisting of the helix of six nucleosomes per turn to form a solenoid with a diameter of 30 nm. In this case, the histone protein can interfere with the transcription of a number of genes and thus regulate their activity.
As a result of the interactions of DNA with histones described above, a segment of a DNA double helix of 186 base pairs with an average diameter of 2 nm and a length of 57 nm is converted into a helix with a diameter of 10 nm and a length of 5 nm. When this helix is subsequently compressed to a fiber with a diameter of 30 nm, the degree of condensation increases another sixfold.
Ultimately, the packaging of a DNA duplex with five histones results in 50-fold condensation of DNA. However, even such a high degree of condensation cannot explain the almost 50,000 - 100,000-fold compaction of DNA in the metaphase chromosome. Unfortunately, the details of further chromatin packaging up to the metaphase chromosome are not yet known, so we can only consider the general features of this process.
Levels of DNA compaction in chromosomes
Each DNA molecule is packaged into a separate chromosome. Human diploid cells contain 46 chromosomes, which are located in the cell nucleus. The total length of the DNA of all chromosomes in a cell is 1.74 m, but the diameter of the nucleus in which the chromosomes are packaged is millions of times smaller. Such compact packaging of DNA in chromosomes and chromosomes in the cell nucleus is ensured by a variety of histone and non-histone proteins that interact in a certain sequence with DNA (see above). Compacting DNA in chromosomes makes it possible to reduce its linear dimensions by approximately 10,000 times - roughly from 5 cm to 5 microns. There are several levels of compaction (Fig. 10).


- DNA double helix is a negatively charged molecule with a diameter of 2 nm and a length of several cm.
- nucleosome level- chromatin looks in an electron microscope as a chain of “beads” - nucleosomes - “on a thread”. The nucleosome is a universal structural unit that is found in both euchromatin and heterochromatin, in the interphase nucleus and metaphase chromosomes.
The nucleosomal level of compaction is ensured by special proteins - histones. Eight positively charged histone domains form the core of the nucleosome around which a negatively charged DNA molecule is wound. This gives a shortening of 7 times, while the diameter increases from 2 to 11 nm.
- solenoid level
The solenoid level of chromosome organization is characterized by twisting of the nucleosome filament and the formation of thicker fibrils 20-35 nm in diameter - solenoids or superbids. The solenoid pitch is 11 nm; there are about 6-10 nucleosomes per turn. Solenoid packing is considered more likely than superbid packing, according to which a chromatin fibril with a diameter of 20-35 nm is a chain of granules, or superbids, each of which consists of eight nucleosomes. At the solenoid level, the linear size of DNA is reduced by 6-10 times, the diameter increases to 30 nm.
- loop level
The loop level is provided by non-histone site-specific DNA-binding proteins that recognize and bind to specific DNA sequences, forming loops of approximately 30-300 kb. The loop ensures gene expression, i.e. the loop is not only a structural, but also a functional formation. Shortening at this level occurs 20-30 times. The diameter increases to 300 nm. Loop-shaped structures such as “lamp brushes” in amphibian oocytes can be seen in cytological preparations. These loops appear to be supercoiled and represent DNA domains, probably corresponding to units of transcription and chromatin replication. Specific proteins fix the bases of the loops and, possibly, some of their internal sections. The loop-like domain organization promotes the folding of chromatin in metaphase chromosomes into helical structures of higher orders.
- domain level
The domain level of chromosome organization has not been studied enough. At this level, the formation of loop domains is noted - structures of threads (fibrils) 25-30 nm thick, which contain 60% protein, 35% DNA and 5% RNA, are practically invisible in all phases of the cell cycle with the exception of mitosis and are somewhat randomly distributed throughout cell nucleus. Loop-shaped structures such as “lamp brushes” in amphibian oocytes can be seen in cytological preparations.
Loop domains are attached at their base to the intranuclear protein matrix in the so-called built-in attachment sites, often referred to as MAR/SAR sequences (MAR, from the English matrix associated region; SAR, from the English scaffold attachment regions) - DNA fragments several hundred in length base pairs that are characterized by a high content (>65%) of A/T nucleotide pairs. Each domain appears to have a single origin of replication and functions as an autonomous superhelical unit. Any loop domain contains many transcription units, the functioning of which is likely coordinated - the entire domain is either in an active or inactive state.
At the domain level, as a result of sequential chromatin packaging, a decrease in the linear dimensions of DNA occurs by approximately 200 times (700 nm).
- chromosomal level
At the chromosomal level, condensation of the prophase chromosome into a metaphase chromosome occurs with compaction of loop domains around the axial framework of non-histone proteins. This supercoiling is accompanied by phosphorylation of all H1 molecules in the cell. As a result, the metaphase chromosome can be depicted as densely packed solenoid loops, coiled into a tight spiral. A typical human chromosome can contain up to 2,600 loops. The thickness of such a structure reaches 1400 nm (two chromatids), and the DNA molecule is shortened by 104 times, i.e. from 5 cm stretched DNA to 5 µm.
Functions of chromosomes
In interaction with extrachromosomal mechanisms, chromosomes provide
- storage of hereditary information
- using this information to create and maintain cellular organization
- regulation of reading hereditary information
- self-duplication of genetic material
- transfer of genetic material from the mother cell to the daughter cells.
There is evidence that when a region of chromatin is activated, i.e. during transcription, first histone H1 and then the histone octet are reversibly removed from it. This causes chromatin decondensation, the sequential transition of a 30-nm chromatin fibril into a 10-nm fibril and its further unfolding into sections of free DNA, i.e. loss of nucleosome structure.
After the discovery of the principle of molecular organization of such a substance as DNA in 1953, molecular biology began to develop. Further in the process of research, scientists found out how DNA is recombined, its composition and how our human genome is structured.
Every day, complex processes occur at the molecular level. How is the DNA molecule structured, what does it consist of? And what role do DNA molecules play in a cell? Let's talk in detail about all the processes occurring inside the double chain.
What is hereditary information?
So where did it all start? Back in 1868 they found it in the nuclei of bacteria. And in 1928, N. Koltsov put forward the theory that all genetic information about a living organism is encrypted in DNA. Then J. Watson and F. Crick found a model for the now well-known DNA helix in 1953, for which they deservedly received recognition and an award - the Nobel Prize.
What is DNA anyway? This substance consists of 2 united threads, or rather spirals. A section of such a chain with certain information is called a gene.

DNA stores all the information about what kind of proteins will be formed and in what order. The DNA macromolecule is a material carrier of incredibly voluminous information, which is recorded in a strict sequence of individual bricks - nucleotides. There are 4 nucleotides in total; they complement each other chemically and geometrically. This principle of complementation, or complementarity, in science will be described later. This rule plays a key role in the encoding and decoding of genetic information.
Since the DNA strand is incredibly long, there are no repetitions in this sequence. Every living creature has its own unique strand of DNA.
Functions of DNA
Functions include storage of hereditary information and its transmission to offspring. Without this function, the genome of a species could not be preserved and developed over thousands of years. Organisms that have undergone severe gene mutations are more likely to not survive or lose the ability to produce offspring. This is how natural protection against the degeneration of the species occurs.

Another essential function is the implementation of stored information. A cell cannot create a single vital protein without those instructions that are stored in a double chain.
Nucleic acid composition
It is now known for certain what the nucleotides themselves—the building blocks of DNA—are made of. They contain 3 substances:
- Orthophosphoric acid.
- Nitrogenous base. Pyrimidine bases - which have only one ring. These include thymine and cytosine. Purine bases, which contain 2 rings. These are guanine and adenine.
- Sucrose. DNA contains deoxyribose, RNA contains ribose.
The number of nucleotides is always equal to the number of nitrogenous bases. In special laboratories, the nucleotide is broken down and the nitrogenous base is isolated from it. This is how the individual properties of these nucleotides and possible mutations in them are studied.
Levels of organization of hereditary information
There are 3 levels of organization: genetic, chromosomal and genomic. All the information needed for the synthesis of a new protein is contained in a small section of the chain - the gene. That is, the gene is considered the lowest and simplest level of information encoding.

Genes, in turn, are assembled into chromosomes. Thanks to this organization of the carrier of hereditary material, groups of characteristics alternate according to certain laws and are transmitted from one generation to another. It should be noted that there are an incredible number of genes in the body, but the information is not lost even when it is recombined many times.
There are several types of genes:
- According to their functional purpose, there are 2 types: structural and regulatory sequences;
- Based on their influence on the processes occurring in the cell, they distinguish: supervital, lethal, conditionally lethal genes, as well as mutator and antimutator genes.
Genes are arranged along the chromosome in a linear order. In chromosomes, information is not focused randomly; there is a certain order. There is even a map that shows the positions, or loci, of genes. For example, it is known that chromosome No. 18 encrypts data about the color of a child’s eyes.
What is a genome? This is the name given to the entire set of nucleotide sequences in an organism’s cell. The genome characterizes an entire species, not an individual.
What is the human genetic code?
The fact is that the entire enormous potential of human development lies already in the period of conception. All hereditary information that is necessary for the development of the zygote and the growth of the child after birth is encrypted in genes. DNA sections are the most basic carriers of hereditary information.
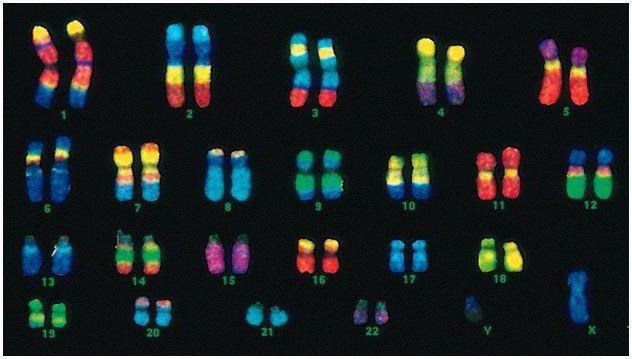
Humans have 46 chromosomes, or 22 somatic pairs plus one sex-determining chromosome from each parent. This diploid set of chromosomes encodes the entire physical appearance of a person, his mental and physical abilities and susceptibility to diseases. Somatic chromosomes are outwardly indistinguishable, but they carry different information, since one of them is from the father, the other from the mother.
The male code differs from the female code in the last pair of chromosomes - XY. The female diploid set is the last pair, XX. Males receive one X chromosome from their biological mother, which is then passed on to their daughters. The sex Y chromosome is passed on to sons.
Human chromosomes vary greatly in size. For example, the smallest pair of chromosomes is No. 17. And the biggest pair is 1 and 3.
The diameter of the double helix in humans is only 2 nm. The DNA is coiled so tightly that it fits inside the small nucleus of a cell, although it would be up to 2 meters long if untwisted. The length of the helix is hundreds of millions of nucleotides.
How is the genetic code transmitted?
So, what role do DNA molecules play in cell division? Genes - carriers of hereditary information - are located inside every cell of the body. To pass on their code to a daughter organism, many creatures divide their DNA into 2 identical helices. This is called replication. During the replication process, DNA unwinds and special “machines” complete each strand. After the genetic helix bifurcates, the nucleus and all organelles begin to divide, and then the entire cell.
But humans have a different process of gene transmission - sexual. The characteristics of the father and mother are mixed, the new genetic code contains information from both parents.
The storage and transmission of hereditary information is possible due to the complex organization of the DNA helix. After all, as we said, the structure of proteins is encrypted in genes. Once created at the time of conception, this code will copy itself throughout life. The karyotype (personal set of chromosomes) does not change during the renewal of organ cells. The transfer of information is carried out with the help of sex gametes - male and female.
Only viruses containing one strand of RNA are not capable of transmitting their information to their offspring. Therefore, they need human or animal cells to reproduce.
Implementation of hereditary information
Important processes constantly occur in the cell nucleus. All information recorded in chromosomes is used to build proteins from amino acids. But the DNA chain never leaves the nucleus, so it needs the help of another important compound: RNA. It is RNA that is able to penetrate the nuclear membrane and interact with the DNA chain.
Through the interaction of DNA and 3 types of RNA, all encoded information is realized. At what level does the implementation of hereditary information occur? All interactions occur at the nucleotide level. Messenger RNA copies a section of the DNA chain and brings this copy to the ribosome. Here the synthesis of a new molecule from nucleotides begins.
In order for the mRNA to copy the necessary part of the chain, the helix unfolds and then, upon completion of the recoding process, is restored again. Moreover, this process can occur simultaneously on 2 sides of 1 chromosome.
Principle of complementarity
They consist of 4 nucleotides - adenine (A), guanine (G), cytosine (C), thymine (T). They are connected by hydrogen bonds according to the rule of complementarity. The work of E. Chargaff helped establish this rule, since the scientist noticed some patterns in the behavior of these substances. E. Chargaff discovered that the molar ratio of adenine to thymine is equal to one. And in the same way, the ratio of guanine to cytosine is always equal to one.
Based on his work, geneticists formed a rule for the interaction of nucleotides. The complementarity rule states that adenine combines only with thymine, and guanine only combines with cytosine. During the decoding of the helix and the synthesis of a new protein in the ribosome, this alternation rule helps to quickly find the necessary amino acid that is attached to the transfer RNA.
RNA and its types
What is hereditary information? nucleotides in a double strand of DNA. What is RNA? What is her job? RNA, or ribonucleic acid, helps extract information from DNA, decode it and, based on the principle of complementarity, create proteins necessary for cells.
There are 3 types of RNA in total. Each of them performs strictly its own function.
- Informational (mRNA), or also called matrix. It goes straight into the center of the cell, into the nucleus. Finds in one of the chromosomes the necessary genetic material to build a protein and copies one of the sides of the double strand. Copying occurs again according to the principle of complementarity.
- Transport is a small molecule that has nucleotide decoders on one side, and amino acids corresponding to the basic code on the other side. The task of tRNA is to deliver it to the “workshop,” that is, to the ribosome, where it synthesizes the necessary amino acid.
- rRNA is ribosomal. It controls the amount of protein that is produced. It consists of 2 parts - an amino acid and a peptide section.
The only difference in decoding is that RNA does not have thymine. Instead of thymine, uracil is present here. But then, during the process of protein synthesis, tRNA still correctly installs all the amino acids. If any failures occur in decoding information, then a mutation occurs.
Repair of damaged DNA molecule
The process of restoring a damaged double strand is called repair. During the repair process, damaged genes are removed.

Then the required sequence of elements is exactly reproduced and cut back into the same place on the chain from where it was removed. All this happens thanks to special chemicals - enzymes.
Why do mutations occur?
Why do some genes begin to mutate and cease to perform their function - storing vital hereditary information? This occurs due to an error in decoding. For example, if adenine is accidentally replaced with thymine.
There are also chromosomal and genomic mutations. Chromosomal mutations occur when sections of hereditary information are lost, duplicated, or even transferred and inserted into another chromosome.

Genomic mutations are the most serious. Their cause is a change in the number of chromosomes. That is, when instead of a pair - a diploid set, a triploid set is present in the karyotype.
The most famous example of a triploid mutation is Down syndrome, in which the personal set of chromosomes is 47. In such children, 3 chromosomes are formed in place of the 21st pair.
There is also a known mutation called polyploidy. But polyploidy occurs only in plants.
The DNA molecule consists of two strands forming a double helix. Its structure was first deciphered by Francis Crick and James Watson in 1953.
At first, the DNA molecule, consisting of a pair of nucleotide chains twisted around each other, gave rise to questions about why it had this particular shape. Scientists call this phenomenon complementarity, which means that only certain nucleotides can be found opposite each other in its strands. For example, adenine is always opposite thymine, and guanine is always opposite cytosine. These nucleotides of the DNA molecule are called complementary.
Schematically it is depicted like this:
T - A
C - G
These pairs form a chemical nucleotide bond, which determines the order of amino acids. In the first case it is a little weaker. The connection between C and G is stronger. Non-complementary nucleotides do not form pairs with each other.
About the building
So, the structure of the DNA molecule is special. It has this shape for a reason: the fact is that the number of nucleotides is very large, and a lot of space is needed to accommodate long chains. It is for this reason that the chains are characterized by a spiral twist. This phenomenon is called spiralization, it allows the threads to shorten by about five to six times.
The body uses some molecules of this type very actively, others rarely. The latter, in addition to spiralization, also undergo such “compact packaging” as superspiralization. And then the length of the DNA molecule decreases by 25-30 times.

What is the “packaging” of a molecule?
The process of supercoiling involves histone proteins. They have the structure and appearance of a spool of thread or a rod. Spiralized threads are wound onto them, which immediately become “compactly packaged” and take up little space. When the need arises to use one or another thread, it is unwound from a spool, for example, a histone protein, and the helix unwinds into two parallel chains. When the DNA molecule is in this state, the necessary genetic data can be read from it. However, there is one condition. Obtaining information is possible only if the structure of the DNA molecule has an untwisted form. Chromosomes that are accessible for reading are called euchromatins, and if they are supercoiled, then they are already heterochromatins.
Nucleic acids
Nucleic acids, like proteins, are biopolymers. The main function is the storage, implementation and transmission of hereditary (genetic information). They come in two types: DNA and RNA (deoxyribonucleic and ribonucleic). The monomers in them are nucleotides, each of which contains a phosphoric acid residue, a five-carbon sugar (deoxyribose/ribose) and a nitrogenous base. The DNA code includes 4 types of nucleotides - adenine (A) / guanine (G) / cytosine (C) / thymine (T). They differ in the nitrogenous base they contain.
In a DNA molecule, the number of nucleotides can be huge - from several thousand to tens and hundreds of millions. Such giant molecules can be examined through an electron microscope. In this case, you will be able to see a double chain of polynucleotide strands, which are connected to each other by hydrogen bonds of the nitrogenous bases of the nucleotides.

Research
During the course of research, scientists discovered that the types of DNA molecules differ in different living organisms. It was also found that guanine of one chain can only bind to cytosine, and thymine to adenine. The arrangement of nucleotides in one chain strictly corresponds to the parallel one. Thanks to this complementarity of polynucleotides, the DNA molecule is capable of doubling and self-reproduction. But first, the complementary chains, under the influence of special enzymes that destroy paired nucleotides, diverge, and then in each of them the synthesis of the missing chain begins. This occurs due to the free nucleotides present in large quantities in each cell. As a result of this, instead of the “mother molecule”, two “daughter” ones are formed, identical in composition and structure, and the DNA code becomes the original one. This process is a precursor to cell division. It ensures the transmission of all hereditary data from mother cells to daughter cells, as well as to all subsequent generations.
How is the gene code read?
Today, not only the mass of a DNA molecule is calculated - it is also possible to find out more complex data that was previously inaccessible to scientists. For example, you can read information about how an organism uses its own cell. Of course, at first this information is in encoded form and has the form of a certain matrix, and therefore it must be transported to a special carrier, which is RNA. Ribonucleic acid is able to penetrate into the cell through the nuclear membrane and read the encoded information inside. Thus, RNA is a carrier of hidden data from the nucleus to the cell, and it differs from DNA in that it contains ribose instead of deoxyribose, and uracil instead of thymine. In addition, RNA is single-stranded.

RNA synthesis
In-depth analysis of DNA has shown that after RNA leaves the nucleus, it enters the cytoplasm, where it can be integrated as a matrix into ribosomes (special enzyme systems). Guided by the information received, they can synthesize the appropriate sequence of protein amino acids. The ribosome learns from the triplet code which type of organic compound needs to be attached to the forming protein chain. Each amino acid has its own specific triplet, which encodes it.
After the formation of the chain is completed, it acquires a specific spatial form and turns into a protein capable of performing its hormonal, construction, enzymatic and other functions. For any organism it is a gene product. It is from it that all kinds of qualities, properties and manifestations of genes are determined.

Genes
Sequencing processes were primarily developed to obtain information about how many genes a DNA molecule has in its structure. And, although research has allowed scientists to make great progress in this matter, it is not yet possible to know their exact number.
Just a few years ago it was assumed that DNA molecules contain approximately 100 thousand genes. A little later, the figure decreased to 80 thousand, and in 1998, geneticists stated that only 50 thousand genes are present in one DNA, which are only 3% of the total DNA length. But the latest conclusions of geneticists were striking. Now they claim that the genome includes 25-40 thousand of these units. It turns out that only 1.5% of chromosomal DNA is responsible for coding proteins.
The research did not stop there. A parallel team of genetic engineering specialists found that the number of genes in one molecule is exactly 32 thousand. As you can see, it is still impossible to get a definitive answer. There are too many contradictions. All researchers rely only on their results.
Was there evolution?
Despite the fact that there is no evidence of the evolution of the molecule (since the structure of the DNA molecule is fragile and small in size), scientists still made one assumption. Based on laboratory data, they voiced the following version: at the initial stage of its appearance, the molecule had the form of a simple self-replicating peptide, which included up to 32 amino acids found in the ancient oceans.
After self-replication, thanks to the forces of natural selection, molecules acquired the ability to protect themselves from external elements. They began to live longer and reproduce in larger quantities. Molecules that found themselves in the lipid bubble had every chance to reproduce themselves. As a result of a series of successive cycles, lipid bubbles acquired the form of cell membranes, and then - the well-known particles. It should be noted that today any section of a DNA molecule is a complex and clearly functioning structure, all the features of which scientists have not yet fully studied.

Modern world
Recently, scientists from Israel have developed a computer that can perform trillions of operations per second. Today it is the fastest car on Earth. The whole secret is that the innovative device is powered by DNA. Professors say that in the near future, such computers will even be able to generate energy.
A year ago, specialists from the Weizmann Institute in Rehovot (Israel) announced the creation of a programmable molecular computing machine consisting of molecules and enzymes. They replaced silicon microchips with them. To date, the team has made further progress. Now just one DNA molecule can provide a computer with the necessary data and the necessary fuel.
Biochemical “nanocomputers” are not a fiction; they already exist in nature and are manifested in every living creature. But often they are not managed by people. A person cannot yet operate on the genome of any plant in order to calculate, say, the number “Pi”.
The idea of using DNA for storing/processing data first came to the minds of scientists in 1994. It was then that a molecule was used to solve a simple mathematical problem. Since then, a number of research groups have proposed various projects related to DNA computers. But here all attempts were based only on the energy molecule. You cannot see such a computer with the naked eye; it looks like a transparent solution of water in a test tube. There are no mechanical parts in it, but only trillions of biomolecular devices - and this is just in one drop of liquid!
Human DNA
People became aware of the type of human DNA in 1953, when scientists were first able to demonstrate to the world a double-stranded DNA model. For this, Kirk and Watson received the Nobel Prize, since this discovery became fundamental in the 20th century.
Over time, of course, they proved that a structured human molecule can look not only like in the proposed version. After conducting a more detailed DNA analysis, they discovered the A-, B- and left-handed form Z-. Form A- is often an exception, since it is formed only if there is a lack of moisture. But this is only possible in laboratory studies; for the natural environment this is anomalous; such a process cannot occur in a living cell.
The B- shape is classic and is known as a double right-handed chain, but the Z- shape is not only twisted in the opposite direction to the left, but also has a more zigzag appearance. Scientists have also identified the G-quadruplex form. Its structure has not 2, but 4 threads. According to geneticists, this form occurs in areas where there is an excess amount of guanine.

Artificial DNA
Today there is already artificial DNA, which is an identical copy of the real one; it perfectly follows the structure of the natural double helix. But, unlike the original polynucleotide, the artificial one has only two additional nucleotides.
Since the dubbing was created based on information obtained from various studies of real DNA, it can also be copied, self-replicating and evolving. Experts have been working on the creation of such an artificial molecule for about 20 years. The result is an amazing invention that can use the genetic code in the same way as natural DNA.
To the four existing nitrogenous bases, geneticists added two additional ones, which were created by chemical modification of natural bases. Unlike natural DNA, artificial DNA turned out to be quite short. It contains only 81 base pairs. However, it also reproduces and evolves.
Replication of a molecule obtained artificially takes place thanks to the polymerase chain reaction, but so far this does not happen independently, but through the intervention of scientists. They independently add the necessary enzymes to the said DNA, placing it in a specially prepared liquid medium.
Final result
The process and final outcome of DNA development can be influenced by various factors, such as mutations. This makes it necessary to study samples of matter so that the analysis result is reliable and reliable. An example is a paternity test. But we can’t help but rejoice that incidents such as mutation are rare. Nevertheless, samples of matter are always rechecked in order to obtain more accurate information based on the analysis.

Plant DNA
Thanks to high sequencing technologies (HTS), a revolution has been made in the field of genomics - DNA extraction from plants is also possible. Of course, obtaining high-quality molecular weight DNA from plant material poses some difficulties due to the large number of copies of mitochondria and chloroplast DNA, as well as the high level of polysaccharides and phenolic compounds. To isolate the structure we are considering in this case, a variety of methods are used.
Hydrogen bond in DNA
The hydrogen bond in the DNA molecule is responsible for the electromagnetic attraction created between a positively charged hydrogen atom that is attached to an electronegative atom. This dipole interaction does not meet the criterion of a chemical bond. But it can occur intermolecularly or in different parts of the molecule, i.e. intramolecularly.

A hydrogen atom attaches to the electronegative atom that is the donor of the bond. An electronegative atom can be nitrogen, fluorine, or oxygen. It - through decentralization - attracts the electron cloud from the hydrogen nucleus to itself and makes the hydrogen atom (partially) positively charged. Since the size of H is small compared to other molecules and atoms, the charge is also small.
DNA decoding
Before deciphering a DNA molecule, scientists first take a huge number of cells. For the most accurate and successful work, about a million of them are needed. The results obtained during the study are constantly compared and recorded. Today, genome decoding is no longer a rarity, but an accessible procedure.

Of course, deciphering the genome of a single cell is an impractical exercise. The data obtained during such studies are of no interest to scientists. But it is important to understand that all currently existing decoding methods, despite their complexity, are not effective enough. They will only allow reading 40-70% of the DNA.
However, Harvard professors recently announced a method through which 90% of the genome can be deciphered. The technique is based on adding primer molecules to isolated cells, with the help of which DNA replication begins. But even this method cannot be considered successful; it still needs to be refined before it can be openly used in science.
