Молекулата на ДНК се наоѓа. ДНК и гени. Нов поглед на ДНК
Биохемиски основи на наследноста.
Генетска улога на нуклеинските киселини.
Нуклеинските киселини се биолошки полимери кои се наоѓаат во сите клетки, од примитивни до сложени. Тие првпат биле откриени од Јохан Фридрих Мишер во 1868 година во клетки богати со нуклеарен материјал (леукоцити, сперма од лосос). Терминот „нуклеински киселини“ беше предложен во 1889 година.
Постојат два вида нуклеински киселини: ДНК, РНК (АТП - мононуклеотид). ДНК и РНК се молекули - матрици. ДНК е содржана околу 6 * 10 -12 g во соматските клетки: во јадрото, митохондриите. РНК е дел од рибозомите и се наоѓа во јадрото и цитоплазмата.
Студијата и доказот за водечката улога на нуклеинските киселини во преносот на наследни информации беа спроведени на вирусни честички. Познато е дека вирусот на мозаикот од тутун е вирулентен за тутунот и хлебните. Вирусната честичка се состои од 95% протеин и 5% нуклеинска киселина. Протеинскиот капсид во вирусните честички беше заменет, но по некое време протеинот во двата вида беше трансформиран во претходната форма.
Кај бактериофагите кои го инфицираат E. coli, протеините на обвивката на фагот беа означени со радиоактивен S, а ДНК на фагот беше означена со радиоактивен P. Во бактериска клетка инфицирана со фагот, беа формирани честички на фагот кои содржеа само радиоактивен P.
Структура и функции на молекулите на ДНК и РНК.
Нуклеинските киселини се биополимери со неправилна структура, чии мономери се нуклеотиди. Нуклеотидот се состои од остатоци од три супстанции: фосфорна киселина, јаглени хидрати - пентоза, азотна база. ДНК нуклеотидите содржат јаглехидрат деоксирибоза, а РНК содржи рибоза. Остатоци од пуринските и пиримидинските азотни бази кои ја сочинуваат ДНК се аденин, гванин, цитозин, тимин. Молекулите на РНК содржат аденин, гванин, цитозин и урацил.
Нуклеотидите се поврзани едни со други преку остаток на фосфорна киселина на еден нуклеотид и јаглени хидрати на друг со силна ковалентна естерска врска, наречена „кислороден мост“. Врската поминува низ петтиот јаглероден атом на јаглехидрат од еден нуклеотид до третиот јаглероден атом на јаглехидрат од друг нуклеотид. Нуклеотидната низа ја претставува примарната структура на нуклеинските киселини. РНК е единечен полинуклеотиден ланец. Структурата на ДНК е двоен полинуклеотиден синџир, намотан во спирала.
Секундарната структура на ДНК се формира кога се појавува втора нишка на ДНК, распоредена според принципот на комплементарност во однос на првата. Вториот синџир е спротивен на првиот (антипаралелен). Азотните основи лежат во рамнина нормална на рамнината на молекулата - ова наликува на спирални скалила. Оградите на ова скалило се остатоци од фосфорна киселина и јаглехидрати, а чекорите се азотни бази.
Азотните бази кои го сочинуваат секој нуклеотид во спротивно насочени синџири се способни да формираат комплементарни водородни врски меѓу себе (поради постојните функционални групи во структурата на секоја азотна база). Аденил нуклеотидот е комплементарен на тимин, гванил на цитозин и обратно. Самите, овие врски се кревки, но молекулата на ДНК „зашиена“ постојано по целата должина со такви врски претставува многу силна врска.
Комплементарност- ова е просторно-структурна и хемиска кореспонденција на азотни бази едни на други; тие се вклопуваат едни со други „како клуч од бравата“.
Една молекула на ДНК може да содржи 108 или повеќе нуклеотиди.
Структурата на молекулата на ДНК како двојна антипаралелна спирала била предложена во 1953 година од американскиот биолог Џејмс Вотсон и англискиот физичар Френсис Крик.
Молекулата на ДНК на кој било жив организам на планетата се состои од само четири типа на нуклеотиди, кои се разликуваат едни од други по азотни бази што ги содржат: аденил, гуанил, тимин и цитозин. Во тоа разноврсност ДНК. Нивната низа е различна, а бројот е бесконечен.
За секој вид на живи организми и за секој организам посебно, оваа низа е индивидуална и строго специфичен .
ОсобеностСтруктурата на ДНК е дека хемиски активните делови на молекулата - азотни бази, се потопуваат во центарот на спиралата и формираат комплементарни врски едни со други, а остатоците од деоксирибозата и фосфорната киселина се наоѓаат на периферијата и го покриваат пристапот до азотни бази. - тие се хемиски неактивни. Оваа структура може да одржува хемиска стабилност долго време. Што друго е потребно за складирање на наследни информации? Токму овие карактеристики на структурата на ДНК ја одредуваат нејзината способност да ги кодира и репродуцира генетските информации.
Силната структура на ДНК е доста тешко да се уништи. Сепак, тоа се случува редовно во клетката - за време на синтезата на РНК и удвојувањето на самата молекула на ДНК пред клеточната делба.
Умножување, репликација на ДНКзапочнува со фактот дека специјален ензим - ДНК полимераза - ја одмотува двојната спирала и ја дели на посебни нишки - се формира вилушка за редупликација. Ензимот делува како брава на патент. На секој синџир со една жица - лепливите краеви на вилушката за редупликација - се синтетизира нов ланец од слободни нуклеотиди во кариоплазмата според принципот на комплементарност. Во две нови молекули на ДНК, едната влакно останува првобитната матична нишка, а втората останува новата ќерка-влакна. Како резултат на тоа, наместо една молекула на ДНК, се појавуваат две молекули со ист точен нуклеотиден состав како и оригиналниот.
Во живите системи се среќаваме со нов тип на реакции непознати во неживата природа. Тие се повикани реакции на синтеза на матрица . Синтезата на матрицата е како фрлање на матрица: новите молекули се синтетизираат во точна согласност со планот поставен во структурата на постоечките молекули. Овие реакции обезбедуваат точна низа на мономерни единици во синтетизираните полимери. Мономерите пристигнуваат на одредена локација на молекулите кои служат како матрица каде што се одвива реакцијата. Доколку таквите реакции се случат како резултат на случајни судири на молекули, тие би се одвивале бескрајно бавно. Синтезата на сложени молекули врз основа на принципот на шаблонот се врши брзо и прецизно со помош на ензими. Синтезата на шаблоните лежи во основата на најважните реакции во синтезата на нуклеинските киселини и протеините. Улогата на матрицата во клетката ја играат молекулите на нуклеинските киселини ДНК или РНК. Мономерните молекули од кои се синтетизира полимерот - нуклеотиди или амино киселини - во согласност со принципот на комплементарност, се лоцирани и фиксирани на матрицата по строго дефиниран редослед. Тогаш мономерните единици се поврзани во полимерен синџир, а готовиот полимер ја напушта матрицата. По ова, матрицата е подготвена за склопување на нова, точно иста молекула на полимер.
Реакциите од типот на матрица се специфична карактеристика на живата клетка. Тие се основата на основното својство на сите живи суштества - способноста да се репродуцира нивниот сопствен вид.
Функции на нуклеинските киселини– складирање и пренос на наследни информации. Молекулите на ДНК кодираат информации за примарната структура на протеинот. Синтезата на мРНК молекули се одвива на ДНК матрицата. Овој процес се нарекува „транскрипција“. И-РНК во процесот на „преведување“ имплементира информации во форма на низа од амино киселини во протеинска молекула.
ДНК на секоја клетка носи информации не само за структурните протеини кои го одредуваат обликот на клетката, туку и за сите ензимски протеини, хормонски протеини и други протеини, како и за структурата на сите видови РНК.
Можеби нуклеинските киселини обезбедуваат различни видови биолошка меморија - имунолошки, невролошка итн., а исто така играат значајна улога во регулирањето на биосинтетичките процеси.
Поврзани информации.
Првиот доказ за улогата на ДНК како носител на наследни информации во организмите привлече огромно внимание на проучувањето на нуклеинските киселини. Во 1869 година, Ф. Мишер изолирал посебна супстанција од клеточните јадра, која ја нарекол нуклеин. По 20 години ова име беше заменето со терминот нуклеинска киселина.Во 1924 година, R. Felgen развил метод за цитолошко препознавање на нуклеинските киселини преку нивното специфично боење и покажал дека ДНК е локализирана во јадрата на клетките, а РНК во цитоплазмата. Во 1936 година А.Н. Белозерски и И.И. Дубровскаја изолирала ДНК во чиста форма од јадрата на растителните клетки. До почетокот на 1930-тите. Беа објаснети основните хемиски принципи на структурата на шеќерите на нуклеинската киселина, а во 1953 година беше создаден структурен модел на ДНК.
Основната структурна единица на нуклеинските киселини е нуклеотид, кој се состои од три хемиски различни делови поврзани со ковалентни врски (сл. 5.2).
Ориз. 5.2. Структурни формули: А- нуклеотиди; б- ДНК; V -РНК (види исто така стр. 110)

Ориз. 5.2. Завршува. Структурни формули: А- нуклеотиди; 6 - ДНК; В- РНК
Првиот дел е шеќер кој содржи пет јаглеродни атоми: деоксирибозаво ДНК и рибозаво РНК.
Вториот дел од нуклеотидот, пуринска или пиримидинска азотна база, е ковалентно поврзан со првиот јаглероден атом на шеќерот, формирајќи структура т.н. нуклеозид.ДНК содржи пурински бази - аденин(А) и гванин(Д) - и пиримидински бази - тимин(Т) и цитозин(В). Соодветните нуклеозиди се нарекуваат деоксиаденозин, деоксигуанозин, деокситимидин и деоксицитидин. РНК ги содржи истите пурински бази како ДНК, пиримидинска база цитозин, а наместо тимин содржи урацил(U); соодветните нуклеозиди се нарекуваат аденозин, гванозин, уридин и цитидин.
Третиот дел од нуклеотидот е фосфатна група, која ги поврзува соседните нуклеозиди во полимерен синџир преку фосфодиестерски врски помеѓу 5-јаглеродниот атом на еден шеќер и 3-јаглеродниот атом на друг (сл. 5.2, б. V). Нуклеотидисе нарекуваат нуклеозиди со една или повеќе фосфатни групи поврзани со естерски врски за 3" или 5 јаглеродни атоми на шеќерот. Синтезата на нуклеотидите претходи на синтезата на нуклеинските киселини; соодветно, нуклеотидите се производи од хемиска или ензимска хидролиза на нуклеинските киселини.
Нуклеинските киселини се многу долги полимерни синџири кои се состојат од мононуклеотиди поврзани со 5- и 3'-фосфодиестерски врски. Непроменета молекула на ДНК содржи, во зависност од видот на организмот, од неколку илјади до многу милиони нуклеотиди, недопрена РНК молекула - од 100 до 100 илјади или повеќе нуклеотиди.
Резултатите од анализите на Е. Чаргаф за нуклеотидниот состав на ДНК од различни видови видови покажаа дека молекуларниот сооднос на различни азотни бази - аденин, гванин, тимин, цитозин - варира во голема мера. Следствено, докажано е дека ДНК воопшто не е монотон полимер кој се состои од идентични тетрануклеотиди, како што се претпоставуваше во 40-тите години. XX век, и дека целосно ја поседува сложеноста неопходна за зачувување и пренос на наследни информации во форма на специфична низа на нуклеотидни бази.
Истражувањето на Е. Чаргаф, исто така, откри карактеристика својствена за сите молекули на ДНК: моларната содржина на аденин е еднаква на содржината на тимин, а моларната содржина на гванин е еднаква на содржината на цитозин. Овие еднаквости се нарекуваат правило за еквивалентност на Чаргаф: [A] = [T], [G] = [C]; бројот на пурините е еднаков на бројот на пиримидините. Во зависност од видот, се менува само односот ([A] + [T])/([G] + [C]) (Табела 5.1).
|
Состав на основи |
Став |
Асиметрија |
|||||
|
причини |
|||||||
|
(A + T)/(G + C) |
|||||||
|
Животни |
|||||||
|
Желка |
|||||||
|
морски рак |
|||||||
|
Морски еж |
|||||||
|
Растенија, печурки |
|||||||
|
пченични никулци |
|||||||
|
Печурка Aspergillus niger |
|||||||
|
Бактерија |
|||||||
|
Ешерихија коли |
|||||||
|
Staphylococcus aureus |
|||||||
|
Clostridium perfringens |
|||||||
|
Брусела абортус |
|||||||
|
Сарцина лутеа |
|||||||
|
Бактериофаги |
|||||||
|
FH 174 (вирусна форма) |
|||||||
|
FH 174 (репликативна форма) |
|||||||
Односот на базите се нарекува нуклеотиден коефициент(вид) специфичност.Откритието на Чаргаф формулирало важна структурна карактеристика на ДНК, која подоцна се рефлектирала во структурниот модел на ДНК од Џ. Вотсон и Ф. Крик (1953), кои всушност покажале дека правилата на Чаргаф не наметнуваат никакви ограничувања за можниот број на комбинации на различни базни секвенци способни да формираат молекули на ДНК.
Концептот на нуклеотидна специфичност ја формираше основата на нова гранка на биологијата - генска систематика, кој функционира со споредување на составот и структурата на нуклеинските киселини за да се изгради природен систем на организми.
Според моделот Вотсон-Крик, молекулата на ДНК се состои од два полинуклеотидни синџири (нишки, нишки), поврзани едни со други со помош на попречни водородни врски помеѓу азотни бази според комплементарниот принцип (аденин од еден синџир е поврзан со две водородни врски со тимин од спротивниот синџир, а гванин и цитозин од различни синџири се поврзани еден со друг со три водородни врски). Во овој случај, два полинуклеотидни синџири од една молекула се анти-паралелни, т.е., спроти 3" крајот на едниот синџир е 5" крајот на другиот синџир и обратно (сл. 5.3). Сепак, треба да се имаат предвид современите податоци дека генетскиот материјал на некои вируси е претставен со едноверижни (едноверижни) молекули на ДНК. Врз основа на рендгенската дифракциона анализа на ДНК, Џ. Вотсон и Ф. Крик, исто така, заклучија дека неговата двоверижна молекула има секундарна структура во форма на спирала извиткана во правец од лево кон десно, која подоцна беше наречена 5. -форма (сл. 5.4). До денес е докажано дека покрај најчестите 5-форми, можно е да се детектираат делови од ДНК кои имаат различна конфигурација - како десничарски (форми А, C), и извртено од десно кон лево (завртено лево, или во облик на Z) (сл. 5.4). Постојат одредени разлики помеѓу овие форми на секундарна структура на ДНК (Табела 5.2). На пример, растојанието помеѓу два соседни пара азотни бази во двоверижна спирала, изразено во нанометри (nm), се карактеризира со различни вредности за 5-форма и Z-форма (0,34 и 0,38 nm, соодветно) . На сл. Слика 5.5 покажува модерни тродимензионални модели на „левак“ и „деснорак“ форми на ДНК.

Ориз. 5.3. шематски приказ на примарната структура на фрагмент од двоверижна молекула на ДНК: А - аденин; G - гванин; Т - тимин; C - цитозин

Ориз. 5.4.

Табела 5.2
Својства на различни форми на ДНК двојни спирали
Молекулите на РНК, во зависност од нивните структурни и функционални карактеристики, се поделени на неколку типови: гласничка РНК (мРНК, или мРНК), рибозомална РНК (рРНК), трансферна РНК (тРНК), мала нуклеарна РНК (снРНК), итн. ДНК, РНК молекулите се секогаш едноверижни (едноверижни). Сепак, тие можат да формираат посложени (секундарни) конфигурации поради комплементарното поврзување на поединечни делови од таков синџир врз основа на интеракцијата на комплементарни азотни бази (A-U и G-C). Како пример, земете ја конфигурацијата на детелина за молекулата на РНК за пренос на фенилаланин (Слика 5.6).

Ориз. 5.6.
Во 1953 година, Д. Вотсон и Ф. Крик предложија модел на структурата на ДНК, кој се базираше на следните постулати:
- 1. ДНК е полимер кој се состои од нуклеотиди поврзани со 3" и 5" фосфодиестерски врски.
- 2. Составот на нуклеотидите на ДНК ги почитува правилата на Чаргаф.
- 3. Молекулата на ДНК има структура со двојна спирала, која потсетува на спирални скалила, што е потврдено со шемите на дифракција на рендгенските зраци на нишките на ДНК кои први ги добиле М. Вилкинс и Р. Френклин.
- 4. Структурата на полимерот, како што е прикажано со киселинско-базната титрација на природната (природна) ДНК, се стабилизира со водородни врски. Титрацијата и загревањето на домашната ДНК предизвикува забележлива промена во нејзините физички својства, особено вискозноста, претворајќи ја во денатурирана форма, а ковалентните врски не се уништуваат.
Нуклеинските киселини се високомолекуларни супстанции кои се состојат од мононуклеотиди, кои се поврзани едни со други во полимерен синџир користејќи 3", 5" фосфодиестерски врски и се пакуваат во клетките на одреден начин.
Нуклеинските киселини се биополимери од два вида: рибонуклеинска киселина (РНК) и деоксирибонуклеинска киселина (ДНК). Секој биополимер се состои од нуклеотиди кои се разликуваат во остатоците од јаглени хидрати (рибоза, деоксирибоза) и една од азотни бази (урацил, тимин). Според овие разлики, нуклеинските киселини го добиле своето име.
Структура на деоксирибонуклеинска киселина
Нуклеинските киселини имаат примарна, секундарна и терцијарна структура.
Примарна структура на ДНК
Примарната структура на ДНК е линеарен полинуклеотиден синџир во кој мононуклеотидите се поврзани со 3", 5" фосфодиестерски врски. Почетен материјал за склопување на синџирот на нуклеинска киселина во клетката е 5"-трифосфатниот нуклеозид, кој како резултат на отстранувањето на остатоците од β и γ фосфорна киселина е способен да го закачи 3" јаглеродниот атом на друг нуклеозид. . Така, 3" јаглеродниот атом на една деоксирибоза е ковалентно поврзан со 5" јаглеродниот атом на друга деоксирибоза преку единечен остаток на фосфорна киселина и формира линеарен полинуклеотиден ланец на нуклеинска киселина. Оттука и името: 3", 5" фосфодиестерски врски. Азотните бази не учествуваат во поврзувањето на нуклеотидите од еден синџир (сл. 1.).
Таквата врска, помеѓу остатоците од молекула на фосфорна киселина на еден нуклеотид и јаглехидратите на друг, доведува до формирање на пентоза-фосфатен скелет на молекулата на полинуклеотид, на кој азотни бази се прикачени на страната една по друга. Нивната низа на распоредување во синџирите на молекули на нуклеинска киселина е строго специфична за клетките на различни организми, т.е. има специфичен карактер (правило на Чаргаф).
Линеарна ДНК синџир, чија должина зависи од бројот на нуклеотиди вклучени во синџирот, има два краја: едниот се нарекува 3" крај и содржи слободен хидроксил, а другиот се нарекува 5" крај и содржи фосфорен киселински остаток. Колото е поларно и може да има правец од 5"->3" и 3"->5". Исклучок е кружната ДНК.
Генетскиот „текст“ на ДНК е составен од код „зборови“ - тројки нуклеотиди наречени кодони. Секциите на ДНК кои содржат информации за примарната структура на сите видови РНК се нарекуваат структурни гени.
Полинуклеотидните ДНК синџири достигнуваат гигантски големини, па на одреден начин се пакуваат во клетката.
Додека го проучувал составот на ДНК, Чаргаф (1949) воспоставил важни обрасци во однос на содржината на поединечните ДНК бази. Тие помогнаа да се открие секундарната структура на ДНК. Овие обрасци се нарекуваат правила на Чаргаф. Правила на Chargaff
Овие правила укажуваат дека при конструирањето на ДНК, мора да се почитува прилично строга кореспонденција (спарување) не на пуринските и пиримидинските бази воопшто, туку конкретно на тимин со аденин и цитозин со гванин. Врз основа на овие правила, во 1953 година, Вотсон и Крик предложија модел на секундарната структура на ДНК, наречен двојна спирала (сл.). |

Секундарна структура на ДНК
Секундарната структура на ДНК е двојна спирала, чиј модел беше предложен од Д. Вотсон и Ф. Крик во 1953 година.
Предуслови за создавање на ДНК модел
Како резултат на првичните анализи, се веруваше дека ДНК од кое било потекло ги содржи сите четири нуклеотиди во еднакви моларни количини. Меѓутоа, во 1940-тите, Е. Чаргаф и неговите колеги, како резултат на анализа на ДНК изолирана од различни организми, јасно покажаа дека тие содржат азотни бази во различни квантитативни соодноси. Чаргаф открил дека иако овие соодноси се исти за ДНК од сите клетки од ист вид на организам, ДНК од различни видови може значително да се разликува во содржината на одредени нуклеотиди. Ова сугерираше дека разликите во односот на азотни бази може да бидат поврзани со некој вид биолошки код. Иако односот на поединечни пурински и пиримидински бази во различни примероци на ДНК се покажа дека е различен, при споредување на резултатите од тестот, се појави одредена шема: во сите примероци, вкупниот број на пурини беше еднаков на вкупниот број на пиримидини (А + G = T + C), количината на аденин беше еднаква на количината на тимин (A = T), а количината на гванин е количината на цитозин (G = C). ДНК изолирана од клетките на цицачите беше генерално побогата со аденин и тимин и релативно посиромашна со гванин и цитозин, додека ДНК од бактерии беше побогата со гванин и цитозин и релативно посиромашна со аденин и тимин. Овие податоци формираа важен дел од фактичкиот материјал врз основа на кој подоцна беше изграден Вотсон-Крик моделот на структурата на ДНК.
Друг важен индиректен показател за можната структура на ДНК беше обезбеден со податоците на Л. Полинг за структурата на протеинските молекули. Полинг покажа дека се можни неколку различни стабилни конфигурации на синџирот на аминокиселини во протеинската молекула. Една вообичаена конфигурација на пептидниот синџир, α-спиралата, е правилна спирална структура. Со оваа структура, можно е формирање на водородни врски помеѓу амино киселините лоцирани на соседните свиоци на синџирот. Полинг ја опиша α-спиралната конфигурација на полипептидниот синџир во 1950 година и сугерираше дека молекулите на ДНК веројатно имаат спирална структура која се одржува на место со водородни врски.
Сепак, највредните информации за структурата на молекулата на ДНК ги дадоа резултатите од анализата на дифракција на Х-зраци. Рендгенските зраци кои минуваат низ ДНК кристал подлежат на дифракција, односно се отклонуваат во одредени насоки. Степенот и природата на отклонувањето на зраците зависат од структурата на самите молекули. Шема на дифракција на Х-зраци (сл. 3) му дава на искусното око голем број индиректни индикации во врска со структурата на молекулите на супстанцијата што се испитува. Анализата на моделите на дифракција на Х-зраците на ДНК доведе до заклучок дека азотните бази (кои имаат рамна форма) се наредени како куп плочи. Шемите на дифракција на Х-зраци открија три главни периоди во структурата на кристалната ДНК: 0,34, 2 и 3,4 nm.

Вотсон-Крик ДНК модел
Врз основа на аналитичките податоци на Чаргаф, моделите на рендген на Вилкинс и истражувањето на хемичарите кои дадоа информации за прецизните растојанија помеѓу атомите во молекулата, аглите помеѓу врските на даден атом и големината на атомите, Вотсон и Крик почна да гради физички модели на поединечните компоненти на молекулата на ДНК на одредено ниво и да ги „приспособува“ еден на друг на таков начин што добиениот систем одговара на различни експериментални податоци [прикажи] .
Уште порано беше познато дека соседните нуклеотиди во синџирот на ДНК се поврзани со фосфодиестерски мостови, поврзувајќи го 5"-јаглеродниот деоксирибозен атом на еден нуклеотид со 3"-јаглеродниот деоксирибозен атом на следниот нуклеотид. Вотсон и Крик не се сомневаа дека периодот од 0,34 nm одговара на растојанието помеѓу последователните нуклеотиди во синџирот на ДНК. Понатаму, може да се претпостави дека периодот од 2 nm одговара на дебелината на ланецот. И со цел да се објасни на која вистинска структура одговара периодот од 3,4 nm, Вотсон и Крик, како и Полинг претходно, предложија дека ланецот е извиткан во форма на спирала (или, поточно, формира спирална линија, бидејќи спирала во строга смисла на овие зборови се добиваат кога намотките формираат конусна, а не цилиндрична површина во просторот). Тогаш период од 3,4 nm ќе одговара на растојанието помеѓу последователните вртења на оваа спирала. Таквата спирала може да биде многу густа или донекаде испружена, односно нејзините вртења можат да бидат рамни или стрмни. Бидејќи периодот од 3,4 nm е точно 10 пати поголем од растојанието помеѓу последователните нуклеотиди (0,34 nm), јасно е дека секое целосно вртење на спиралата содржи 10 нуклеотиди. Од овие податоци, Вотсон и Крик можеа да ја пресметаат густината на полинуклеотидниот синџир извиткан во спирала со дијаметар од 2 nm, со растојание помеѓу свиоците од 3,4 nm. Се испостави дека таков синџир ќе има густина што е половина од вистинската густина на ДНК, што веќе беше познато. Морав да претпоставам дека молекулата на ДНК се состои од два синџири - дека е двојна спирала од нуклеотиди.
Следната задача беше, се разбира, да се разјаснат просторните односи помеѓу двата синџири што ја формираат двојната спирала. Испробајќи голем број опции за распоредот на синџирите на нивниот физички модел, Вотсон и Крик открија дека сите достапни податоци најдобро се совпаѓаат со опцијата во која два полинуклеотидни спирали одат во спротивни насоки; во овој случај, синџирите што се состојат од остатоци од шеќер и фосфат ја формираат површината на двојната спирала, а пурините и пиримидините се наоѓаат внатре. Основите лоцирани една спроти друга, кои припаѓаат на два синџири, се поврзани во парови со водородни врски; Токму овие водородни врски ги држат синџирите заедно, со што се фиксира целокупната конфигурација на молекулата.
Двојната спирала на ДНК може да се замисли како скала од јаже што е извиткана на спирален начин, така што нејзините скали остануваат хоризонтални. Тогаш двете надолжни јажиња ќе одговараат на синџири од шеќер и фосфатни остатоци, а попречните шипки ќе одговараат на парови азотни бази поврзани со водородни врски.
Како резултат на понатамошно проучување на можните модели, Вотсон и Крик заклучиле дека секоја „прекрстена шипка“ треба да се состои од еден пурин и еден пиримидин; во период од 2 nm (што одговара на дијаметарот на двојната спирала), нема да има доволно простор за два пурина, а двата пиримидини не би можеле да бидат доволно блиску еден до друг за да формираат соодветни водородни врски. Продлабочената студија на деталниот модел покажа дека аденинот и цитозинот, додека формираат комбинација со соодветна големина, сè уште не може да се позиционираат на таков начин што меѓу нив би се формирале водородни врски. Слични извештаи принудија да се исклучи комбинацијата гванин - тимин, додека комбинациите аденин - тимин и гванин - цитозин се покажаа како сосема прифатливи. Природата на водородните врски е таква што аденин формира пар со тимин, а гванин со цитозин. Оваа идеја за специфично спарување на базите овозможи да се објасни „правилото Чаргаф“, според кое во која било молекула на ДНК, количината на аденин е секогаш еднаква на содржината на тимин, а количината на гванин е секогаш еднаква на количината. на цитозин. Две водородни врски се формираат помеѓу аденин и тимин, а три помеѓу гванин и цитозин. Поради оваа специфичност, формирањето на водородни врски против секој аденин во еден синџир предизвикува формирање на тимин од другиот; на ист начин, само цитозин може да биде спроти секој гванин. Така, синџирите се комплементарни едни со други, односно низата на нуклеотиди во еден синџир уникатно ја одредува нивната низа во другиот. Двата синџири се движат во спротивни насоки и нивните терминални фосфатни групи се на спротивните краеви на двојната спирала.
Како резултат на нивното истражување, во 1953 година Вотсон и Крик предложија модел на структурата на молекулата на ДНК (сл. 3), кој останува релевантен до денес. Според моделот, молекулата на ДНК се состои од два комплементарни полинуклеотидни синџири. Секоја нишка на ДНК е полинуклеотид кој се состои од неколку десетици илјади нуклеотиди. Во него, соседните нуклеотиди формираат правилен столб на пентоза-фосфат поради поврзување на остаток на фосфорна киселина и деоксирибоза со силна ковалентна врска. Азотните бази на едниот полинуклеотиден синџир се распоредени во строго дефиниран редослед наспроти азотните бази на другиот. Алтернацијата на азотни бази во полинуклеотидниот синџир е неправилна.
Распоредот на азотни бази во синџирот на ДНК е комплементарен (од грчкиот „комплемент“ - додаток), т.е. Тиминот (Т) е секогаш против аденин (А), а само цитозинот (C) е против гванин (G). Ова се објаснува со фактот дека A и T, како и G и C, строго одговараат еден на друг, т.е. се надополнуваат еден со друг. Оваа кореспонденција е одредена од хемиската структура на базите, што овозможува формирање на водородни врски во пуринскиот и пиримидинскиот пар. Постојат две врски помеѓу А и Т и три меѓу G и C. Овие врски обезбедуваат делумна стабилизација на молекулата на ДНК во вселената. Стабилноста на двојната спирала е директно пропорционална со бројот на G≡C врски, кои се постабилни во споредба со врските A=T.
Познатата низа на распоред на нуклеотиди во еден синџир на ДНК овозможува, според принципот на комплементарност, да се воспостават нуклеотиди на друг синџир.
Дополнително, утврдено е дека азотни бази кои имаат ароматична структура во воден раствор се наоѓаат една над друга, формирајќи, како да се, куп монети. Овој процес на формирање купишта органски молекули се нарекува редење. Полинуклеотидните синџири на молекулата на ДНК на моделот Вотсон-Крик што се разгледува имаат слична физичко-хемиска состојба, нивните азотни бази се наредени во форма на куп монети, меѓу чии рамнини се појавуваат интеракции на ван дер Валс (редење интеракции).
Водородните врски помеѓу комплементарни бази (хоризонтално) и интеракциите на натрупување помеѓу рамнините на базите во полинуклеотидниот синџир поради ван дер Валсовите сили (вертикално) обезбедуваат на молекулата на ДНК дополнителна стабилизација во просторот.
Шеќерните фосфатни столбови на двата синџири се свртени кон надвор, а основите се свртени навнатре, една кон друга. Насоката на синџирите во ДНК е антипаралелна (едниот од нив има насока од 5"->3", другиот - 3"->5", т.е. 3" крајот на едниот синџир се наоѓа спроти 5" крајот на другиот.). Синџирите формираат десничарски спирали со заедничка оска. Едно вртење на спиралата е 10 нуклеотиди, големината на кривината е 3,4 nm, висината на секој нуклеотид е 0,34 nm, дијаметарот на спиралата е 2,0 nm. Како резултат на ротација на една нишка околу друга, се формираат голем жлеб (околу 20 А во дијаметар) и помал жлеб (околу 12 А во дијаметар) на двојната спирала на ДНК. Оваа форма на двојната спирала Вотсон-Крик подоцна била наречена Б-форма. Во клетките, ДНК обично постои во Б форма, која е најстабилна.
Функции на ДНК
Предложениот модел објасни многу биолошки својства на деоксирибонуклеинската киселина, вклучувајќи складирање на генетски информации и разновидност на гени обезбедени од широк спектар на секвенцијални комбинации од 4 нуклеотиди и фактот за постоење на генетски код, способност за само-репродукција и пренесува генетски информации обезбедени од процесот на репликација и имплементација на генетски информации во форма на протеини, како и сите други соединенија формирани со помош на ензимски протеини.
Основни функции на ДНК.
- ДНК е носител на генетски информации, што е обезбедено со фактот на постоење на генетски код.
- Репродукција и пренос на генетски информации низ генерации клетки и организми. Оваа функционалност е обезбедена со процесот на репликација.
- Имплементација на генетски информации во форма на протеини, како и сите други соединенија формирани со помош на ензимски протеини. Оваа функција е обезбедена од процесите на транскрипција и превод.

Форми на организација на двоверижна ДНК
ДНК може да формира неколку типови на двојни спирали (сл. 4). Во моментов, веќе се познати шест форми (од A до E и Z-форма).
Структурните форми на ДНК, како што утврди Розалинд Френклин, зависат од заситеноста на молекулата на нуклеинската киселина со вода. Во студиите на ДНК влакна користејќи анализа на дифракција на Х-зраци, се покажа дека моделот на Х-зраци радикално зависи од релативната влажност на кој степен на заситеност со вода на ова влакно се одвива експериментот. Ако влакното било доволно заситено со вода, тогаш се добива една радиографија. Кога се исуши, се појави сосема поинаква шема на рендген, многу различна од моделот на рендген на влакна со висока влажност.
Молекулата на ДНК со висока влажност се нарекува Б-форма. Под физиолошки услови (ниска концентрација на сол, висок степен на хидратација), доминантен структурен тип на ДНК е Б-формата (главната форма на двоверижна ДНК - моделот Вотсон-Крик). Чекорот на спиралата на таквата молекула е 3,4 nm. Има 10 комплементарни парови по кривина во форма на изопачени купишта „монети“ - азотни основи. Купиштата се држат заедно со водородни врски помеѓу две спротивставени „монети“ од стеговите и се „намотани“ со две ленти од фосфодиестерски рбет извиткани во десна спирала. Рамнините на азотните основи се нормални на оската на спиралата. Соседните комплементарни парови се ротираат еден во однос на друг за 36°. Дијаметарот на спиралата е 20 А, при што пуринскиот нуклеотид зафаќа 12 А, а пиримидинскиот нуклеотид 8 А.
Молекулата на ДНК со помала влажност се нарекува А-форма. А-формата се формира во услови на помалку висока хидратација и при поголема содржина на јони Na + или K +. Оваа поширока спирална конформација на десната страна има 11 базни парови на вртење. Рамнините на азотните основи имаат посилен наклон кон оската на спиралата, тие се отстапени од нормалата кон оската на спиралата за 20°. Ова подразбира присуство на внатрешна празнина со дијаметар од 5А. Растојанието помеѓу соседните нуклеотиди е 0,23 nm, должината на вртењето е 2,5 nm, а дијаметарот на спиралата е 2,3 nm.
Првично се сметаше дека формата А на ДНК е помалку важна. Меѓутоа, подоцна стана јасно дека А-формата на ДНК, како и Б-формата, има огромно биолошко значење. Спиралата на РНК-ДНК во комплексот шаблон-прајмер има А-форма, како и структури на спирала РНК-РНК и РНК фиба (2'-хидроксилната група на рибоза ги спречува молекулите на РНК да формираат Б-форма). А-формата на ДНК се наоѓа во спорите. Утврдено е дека А-формата на ДНК е 10 пати поотпорна на УВ зраците од Б-формата.
А-формата и Б-формата се нарекуваат канонски форми на ДНК.
Обрасци C-Eисто така деснак, нивното формирање може да се забележи само во специјални експерименти и, очигледно, тие не постојат in vivo. Ц формата на ДНК има структура слична на Б ДНК. Бројот на базни парови по вртење е 9,33, должината на вртењето на спиралата е 3,1 nm. Основните парови се наклонети под агол од 8 степени во однос на нормалната положба на оската. Жлебовите се слични по големина на жлебовите на Б-ДНК. Во овој случај, главниот жлеб е нешто поплиток, а помалиот жлеб е подлабок. Природните и синтетичките ДНК полинуклеотиди може да се трансформираат во C-форма.
| Табела 1. Карактеристики на некои типови на структури на ДНК | |||
| Спирален тип | А | Б | З |
| Спирален терен | 0,32 nm | 3,38 nm | 4,46 nm |
| Спирален пресврт | Во право | Во право | Лево |
| Број на парови на бази по вртење | 11 | 10 | 12 |
| Растојание помеѓу базните рамнини | 0,256 nm | 0,338 nm | 0,371 nm |
| Конформација на гликозидна врска | анти | анти | анти-Ц грев-Г |
| Конформација на прстенот на фуранозата | C3"-ендо | C2"-ендо | C3"-ендо-Г C2"-ендо-C |
| Ширина на жлебот, мал/голем | 1,11/0,22 nm | 0,57/1,17 nm | 0,2/0,88 nm |
| Длабочина на жлебот, мал/голем | 0,26/1,30 nm | 0,82/0,85 nm | 1,38/0,37 nm |
| Дијаметар на спирала | 2,3 nm | 2,0 nm | 1,8 nm |
Структурни елементи на ДНК
(не-канонски ДНК структури)
Структурните елементи на ДНК вклучуваат необични структури ограничени со некои посебни секвенци:
|

ДНК во форма на Zбеше откриен во 1979 година додека го проучуваше хексануклеотидот d(CG)3 -. Го откриле професорот на МИТ Александар Рич и неговите колеги. З-формата стана еден од најважните структурни елементи на ДНК поради фактот што нејзиното формирање е забележано во областите на ДНК каде пурините се менуваат со пиримидини (на пример, 5'-GCGCGC-3'), или во повторувања 5 '-CGCGCG-3' што содржи метилиран цитозин. Суштински услов за формирање и стабилизација на Z-DNA беше присуството на пурински нуклеотиди во него во син конформацијата, наизменично со пиримидинските бази во анти-конформацијата.
Природните молекули на ДНК главно постојат во десна Б-форма освен ако не содржат секвенци како (CG)n. Меѓутоа, ако таквите секвенци се дел од ДНК, тогаш овие делови, кога се менува јонската јачина на растворот или катјоните кои го неутрализираат негативниот полнеж на рамката на фосфодиестер, овие делови може да се трансформираат во Z-форма, додека другите делови на ДНК во синџирот останува во класичната Б-форма. Можноста за таква транзиција укажува на тоа дека двете нишки во двојната спирала на ДНК се во динамична состојба и можат да се одмотаат релативно една на друга, движејќи се од десната во левата и обратно. Биолошките последици од таквата лабилност, која овозможува конформациски трансформации на структурата на ДНК, сè уште не се целосно разбрани. Се верува дека делови од Z-DNA играат одредена улога во регулирањето на експресијата на одредени гени и учествуваат во генетската рекомбинација.
З-формата на ДНК е лева двојна спирала во која фосфодиестерскиот столб се наоѓа во цик-цак шема по должината на оската на молекулата. Оттука и името на молекулата (цик-цак)-DNK. З-ДНК е најмалку извиткана (12 базни парови на вртење) и најтенка ДНК позната во природата. Растојанието помеѓу соседните нуклеотиди е 0,38 nm, должината на вртењето е 4,56 nm, а дијаметарот на Z-DNA е 1,8 nm. Покрај тоа, појавата на оваа молекула на ДНК се одликува со присуство на еден жлеб.
З формата на ДНК е пронајдена во прокариотски и еукариотски клетки. Сега се добиени антитела кои можат да ја разликуваат Z-формата од B-формата на ДНК. Овие антитела се врзуваат за одредени региони на џиновските хромозоми на клетките на плунковните жлезди на Drosophila (Dr. melanogaster). Реакцијата на врзување е лесно да се следи поради необичната структура на овие хромозоми, во кои погустите региони (дискови) се контрастни со помалку густите региони (меѓудискови). З-ДНК регионите се наоѓаат во меѓудисковите. Од ова произлегува дека Z-формата всушност постои во природни услови, иако големини на одделни делови од Z-формата сè уште се непознати.

(инвертори) се најпознатите и најчести базни секвенци во ДНК. Палиндром е збор или фраза што се чита исто од лево кон десно и обратно. Примери за такви зборови или фрази се: КОЛИБА, КОЗАК, ПОПЛАВА И РОЗАТА ПАДНА НА АЗОРОВАТА ШАПА. Кога се применува на делови од ДНК, овој термин (палиндром) значи иста алтернација на нуклеотиди по должината на синџирот од десно кон лево и лево кон десно (како буквите во зборот „колиба“ итн.).
Палиндромот се карактеризира со присуство на превртени повторувања на базните секвенци кои имаат симетрија од втор ред во однос на две нишки на ДНК. Ваквите секвенци, од очигледни причини, се самокомплементарни и имаат тенденција да формираат фиба или крстовидни структури (сл.). Шипките за коса им помагаат на регулаторните протеини да препознаат каде е копиран генетскиот текст на ДНК на хромозомот.
Кога е присутно превртено повторување на истата нишка на ДНК, секвенцата се нарекува повторување на огледалото. Повторувањата на огледалото немаат својства на самокомплементарност и, според тоа, не се способни да формираат фиба или крстовидни структури. Секвенците од овој тип се наоѓаат во речиси сите големи молекули на ДНК и може да се движат од само неколку базни парови до неколку илјади базни парови.
Присуството на палиндроми во форма на крстовидни структури во еукариотските клетки не е докажано, иако одреден број на крстовидни структури се откриени in vivo во клетките на E. coli. Присуството на самокомплементарни секвенци во РНК или едноверижна ДНК е главната причина за превиткување на синџирот на нуклеинска киселина во раствори во одредена просторна структура, која се карактеризира со формирање на многу „шноли“.

ДНК од H-формае спирала формирана од три ДНК нишки - ДНК тројна спирала. Тоа е комплекс од двојна спирала Вотсон-Крик со трета едноверижна ДНК влакно, која се вклопува во нејзиниот главен жлеб, формирајќи таканаречен пар Хугстин.
Формирањето на таков триплекс се јавува како резултат на превиткување на двојна спирала на ДНК на таков начин што половина од нејзиниот пресек останува во форма на двојна спирала, а другата половина е одвоена. Во овој случај, еден од исклучените спирали формира нова структура со првата половина од двојната спирала - тројна спирала, а втората се покажува како неструктурирана, во форма на едножичен пресек. Карактеристика на оваа структурна транзиција е нејзината остра зависност од pH вредноста на медиумот, чии протони ја стабилизираат новата структура. Поради оваа карактеристика, новата структура беше наречена H-форма на ДНК, чиешто формирање беше откриено во супернамотани плазмиди кои содржат хомопурин-хомопиримидински региони, кои се повторување на огледалото.
Во понатамошни студии, беше утврдено дека е можно да се изврши структурна транзиција на некои двоверижни полинуклеотиди хомопурин-хомопиримидин со формирање на триверижна структура која содржи:
- една хомопурин и две хомопиримидински нишки ( Триплекс Py-Pu-Py) [Хугстин интеракција].
Составните блокови на триплексот Py-Pu-Py се канонски изоморфни CGC+ и TAT тријади. За стабилизација на триплексот е потребна протонација на тријадата CGC+, така што овие триплекси зависат од рН на растворот.
- една хомопиримидин и две хомопурин нишки ( Триплекс Пи-Пу-Пу) [инверзна Хугстин интеракција].
Составните блокови на триплексот Py-Pu-Pu се канонски изоморфни CGG и TAA тријади. Суштинско својство на триплексите Py-Pu-Pu е зависноста на нивната стабилност од присуството на двојно наелектризирани јони, а потребни се различни јони за да се стабилизираат триплексите од различни секвенци. Бидејќи формирањето на триплекси Py-Pu-Pu не бара протонирање на нивните составни нуклеотиди, таквите триплекси можат да постојат на неутрална pH вредност.
Забелешка: директните и обратните интеракции на Хогстин се објаснуваат со симетријата на 1-метилтимин: ротација од 180° резултира со тоа што атомот О2 го зазема местото на атомот О4, додека системот на водородни врски е зачуван.
Познати се два вида тројни спирали:
- паралелни тројни спирали во кои поларитетот на третото влакно се совпаѓа со поларитетот на хомопуринскиот синџир на дуплексот Вотсон-Крик
- антипаралелни тројни спирали, во кои поларитетите на третиот и хомопуринскиот синџир се спротивни.

Г-квадриплекс- ДНК со 4 верига. Оваа структура се формира ако има четири гванини, кои го формираат таканаречениот Г-квадруплекс - тркалезен танц од четири гванини.
Првите навестувања за можноста за формирање на такви структури беа примени долго пред пробивот на работата на Вотсон и Крик - уште во 1910 година. Тогаш германскиот хемичар Ивар Банг открил дека една од компонентите на ДНК - гуанозинска киселина - формира гелови во високи концентрации, додека другите компоненти на ДНК го немаат ова својство.
Во 1962 година, со помош на методот на дифракција на Х-зраци, беше можно да се утврди структурата на клетките на овој гел. Се покажа дека е составен од четири остатоци од гванин, кои се поврзуваат едни со други во круг и формираат карактеристичен квадрат. Во центарот, врската е поддржана од метален јон (Na, K, Mg). Истите структури можат да се формираат во ДНК ако содржи многу гванин. Овие рамни квадрати (Г-квартети) се наредени за да формираат прилично стабилни, густи структури (Г-четворица).
Четири одделни нишки на ДНК може да се вткаат во комплекси со четири вериги, но ова е прилично исклучок. Почесто, една нишка на нуклеинска киселина е едноставно врзана во јазол, формирајќи карактеристични згуснувања (на пример, на краевите на хромозомите), или двоверижна ДНК во некои региони богати со гванин формира локален квадриплекс.
Најмногу е проучено постоењето на квадриплекси на краевите на хромозомите - кај теломерите и кај промоторите на туморот. Сепак, сè уште не е позната целосна слика за локализацијата на таквата ДНК во човечките хромозоми.
Сите овие необични ДНК структури во линеарна форма се нестабилни во споредба со ДНК од Б-форма. Сепак, ДНК често постои во кружна форма на тополошка напнатост кога има нешто што се нарекува суперсвиткување. Под овие услови, лесно се формираат неканонски структури на ДНК: Z-форми, „крстови“ и „шноли“, H-форми, квадриплекси на гванин и i-мотив.
- Превиткана форма - забележана кога се ослободува од клеточното јадро без да се оштети рбетот на пентоза фосфат. Има форма на супер-извртени затворени прстени. Во суперсвиткана состојба, двојната спирала на ДНК е „извртена на себе“ барем еднаш, односно содржи најмалку едно превртување (има облик на фигура осум).
- Опуштена состојба на ДНК - забележана со еден прекин (прекин на една жичка). Во овој случај, суперспиралите исчезнуваат и ДНК добива форма на затворен прстен.
- Линеарната форма на ДНК се забележува кога две нишки од двојна спирала се скршени.
Терциерна структура на ДНК
Терциерна структура на ДНКсе формира како резултат на дополнително извртување во просторот на молекула со двојна спирала - нејзино превиткување. Суперсвиткување на молекулата на ДНК во еукариотските клетки, за разлика од прокариотите, се јавува во форма на комплекси со протеини.
Речиси целата ДНК на еукариотите се наоѓа во хромозомите на јадрата; само мала количина е содржана во митохондриите и во растенијата во пластидите. Главната супстанција на хромозомите на еукариотските клетки (вклучувајќи ги и човечките хромозоми) е хроматин, кој се состои од двоверижна ДНК, хистон и нехистонски протеини.
Хистонски хроматин протеини
Хистоните се едноставни протеини кои сочинуваат до 50% од хроматинот. Во сите проучувани животински и растителни клетки, пронајдени се пет главни класи на хистони: H1, H2A, H2B, H3, H4, кои се разликуваат по големина, состав на аминокиселини и полнеж (секогаш позитивни).
Хистон H1 од цицачи се состои од единечен полипептиден ланец кој содржи приближно 215 амино киселини; големините на другите хистони варираат од 100 до 135 аминокиселини. Сите тие се спирализирани и искривени во глобула со дијаметар од околу 2,5 nm и содржат невообичаено голема количина на позитивно наелектризирани амино киселини лизин и аргинин. Хистоните можат да бидат ацетилирани, метилирани, фосфорилирани, поли(ADP)-рибозилирани, а хистоните H2A и H2B се ковалентно поврзани со убиквитин. Улогата на таквите модификации во формирањето на структурата и извршувањето на функциите од страна на хистоните сè уште не е целосно разјаснета. Се претпоставува дека тоа е нивната способност да комуницираат со ДНК и да обезбедат еден од механизмите за регулирање на дејството на гените.
Хистоните комуницираат со ДНК првенствено преку јонски врски (солни мостови) формирани помеѓу негативно наелектризираните фосфатни групи на ДНК и позитивно наелектризираните лизински и аргинински остатоци од хистоните.
Не-хистонски хроматински протеини
Нехистонските протеини, за разлика од хистоните, се многу разновидни. Изолирани се до 590 различни фракции на нехистонски протеини кои се врзуваат за ДНК. Тие се нарекуваат и кисели протеини, бидејќи нивната структура е доминирана од кисели амино киселини (тие се полианиони). Разновидноста на нехистонските протеини е поврзана со специфична регулација на активноста на хроматин. На пример, ензимите потребни за репликација и експресија на ДНК може минливо да се врзат за хроматинот. Други протеини, да речеме, оние кои се вклучени во различни регулаторни процеси, се врзуваат за ДНК само во одредени ткива или во одредени фази на диференцијација. Секој протеин е комплементарен на одредена секвенца на нуклеотиди на ДНК (локација на ДНК). Оваа група вклучува:
- фамилија на специфични за локацијата цинк-прст протеини. Секој „цинк прст“ препознава одредена локација која се состои од 5 нуклеотидни парови.
- фамилија на локациски специфични протеини - хомодимери. Фрагментот од таков протеин во контакт со ДНК има структура на спирала-сврт-спирала.
- гел протеини со висока подвижност (HMG протеини) се група на структурни и регулаторни протеини кои постојано се поврзуваат со хроматин. Тие имаат молекуларна тежина помала од 30 kDa и се карактеризираат со висока содржина на наелектризирани амино киселини. Поради нивната мала молекуларна тежина, HMG протеините имаат висока мобилност за време на електрофорезата на полиакриламид гел.
- ензими за репликација, транскрипција и поправка.
Со учество на структурни, регулаторни протеини и ензими вклучени во синтезата на ДНК и РНК, нуклеозомската нишка се претвора во високо кондензиран комплекс на протеини и ДНК. Добиената структура е 10.000 пати пократка од оригиналната молекула на ДНК.

Хроматин
Хроматинот е комплекс од протеини со нуклеарна ДНК и неоргански материи. Најголемиот дел од хроматинот е неактивен. Содржи цврсто спакувана, кондензирана ДНК. Ова е хетерохроматин. Постојат конститутивен, генетски неактивен хроматин (сателитска ДНК) кој се состои од неизразени региони, и факултативен - неактивен во голем број генерации, но под одредени околности способен за изразување.
Активниот хроматин (еухроматин) е некондензиран, т.е. спакувани помалку цврсто. Во различни ќелии неговата содржина се движи од 2 до 11%. Во мозочните клетки е најзастапен - 10-11%, во клетките на црниот дроб - 3-4 и бубрежните клетки - 2-3%. Забележана е активна транскрипција на еухроматин. Покрај тоа, неговата структурна организација дозволува истите генетски информации за ДНК својствени за даден тип на организам да се користат различно во специјализирани клетки.
Во електронски микроскоп, сликата на хроматин наликува на зрнца: сферични задебелувања со големина од околу 10 nm, разделени со мостови слични на нишки. Овие сферични задебелувања се нарекуваат нуклеозоми. Нуклеозомот е структурна единица на хроматин. Секој нуклеозом содржи 146-bp превиткан ДНК сегмент ранет за да формира 1,75 вртења налево по нуклеозомско јадро. Нуклеозомското јадро е хистонски октамер кој се состои од хистони H2A, H2B, H3 и H4, по две молекули од секој тип (сл. 9), кој изгледа како диск со дијаметар од 11 nm и дебелина од 5,7 nm. Петтиот хистон, H1, не е дел од нуклеозомалното јадро и не е вклучен во процесот на намотување на ДНК на хистонскиот октамер. Тој контактира со ДНК на местата каде што двојната спирала влегува и излегува од нуклеозомалното јадро. Станува збор за интеркор (поврзувачки) делови на ДНК, чија должина варира во зависност од типот на клетката од 40 до 50 нуклеотидни парови. Како резултат на тоа, должината на фрагментот на ДНК вклучен во нуклеозомите исто така варира (од 186 до 196 нуклеотидни парови).
Нуклеозомите содржат приближно 90% ДНК, а остатокот се поврзувачи. Се верува дека нуклеозомите се фрагменти од „тивок“ хроматин, а поврзувачот е активен. Сепак, нуклеозомите можат да се расплетуваат и да станат линеарни. Расклопените нуклеозоми се веќе активен хроматин. Ова јасно ја покажува зависноста на функцијата од структурата. Може да се претпостави дека колку повеќе хроматин е содржан во глобуларните нуклеозоми, толку е помалку активен. Очигледно, во различни клетки нееднаков дел од хроматинот во мирување е поврзан со бројот на такви нуклеозоми.
На електронски микроскопски фотографии, во зависност од условите на изолација и степенот на истегнување, хроматин може да изгледа не само како долга нишка со задебелување - „монистра“ на нуклеозоми, туку и како пократок и погуст фибрил (влакна) со дијаметар од 30 nm. Во овој случај, хистонскиот протеин може да се меша со транскрипцијата на голем број гени и на тој начин да ја регулира нивната активност.
Како резултат на интеракциите на ДНК со хистоните опишани погоре, сегмент од двојна спирала на ДНК од 186 базни парови со просечен дијаметар од 2 nm и должина од 57 nm се претвора во спирала со дијаметар од 10 nm и должина од 5 nm. Кога оваа спирала последователно ќе се компресира на влакно со дијаметар од 30 nm, степенот на кондензација се зголемува уште шест пати.
На крајот на краиштата, пакувањето на ДНК дуплекс со пет хистони резултира со 50-кратна кондензација на ДНК. Сепак, дури и таков висок степен на кондензација не може да го објасни речиси 50.000 - 100.000-кратното набивање на ДНК во метафазниот хромозом. За жал, деталите за понатамошното пакување на хроматин до метафазниот хромозом сè уште не се познати, така што можеме да ги разгледаме само општите карактеристики на овој процес.
Нивоа на набивање на ДНК во хромозомите
Секоја молекула на ДНК е спакувана во посебен хромозом. Човечките диплоидни клетки содржат 46 хромозоми, кои се наоѓаат во клеточното јадро. Вкупната должина на ДНК на сите хромозоми во клетката е 1,74 m, но дијаметарот на јадрото во кое се спакувани хромозомите е милиони пати помал. Таквото компактно пакување на ДНК во хромозомите и хромозомите во клеточното јадро е обезбедено со различни хистонски и нехистонски протеини кои комуницираат во одредена низа со ДНК (види погоре). Набивањето на ДНК во хромозомите овозможува да се намалат нејзините линеарни димензии за приближно 10.000 пати - приближно од 5 см до 5 микрони. Постојат неколку нивоа на набивање (сл. 10).


- Двојната спирала на ДНК е негативно наелектризирана молекула со дијаметар од 2 nm и должина од неколку см.
- ниво на нуклеозом- хроматинот изгледа во електронски микроскоп како синџир од „монистра“ - нуклеозоми - „на конец“. Нуклеозомот е универзална структурна единица која се наоѓа и во ехроматин и во хетерохроматин, во меѓуфазното јадро и во метафазните хромозоми.
Нуклеозомското ниво на набивање е обезбедено со специјални протеини - хистони. Осум позитивно наелектризирани хистонски домени го формираат јадрото на нуклеозомот околу кој е намотана негативно наелектризираната молекула на ДНК. Ова дава скратување од 7 пати, додека дијаметарот се зголемува од 2 до 11 nm.
- ниво на електромагнет
Соленоидното ниво на организација на хромозомот се карактеризира со извртување на нуклеозомската нишка и формирање на подебели фибрили со дијаметар од 20-35 nm - соленоиди или супербиди. Теренот на соленоидот е 11 nm; има околу 6-10 нуклеозоми на вртење. Соленоидното пакување се смета за поверојатно од супербидното пакување, според кое хроматинскиот фибрил со дијаметар од 20-35 nm е синџир од гранули или супербиди, од кои секоја се состои од осум нуклеозоми. На ниво на соленоид, линеарната големина на ДНК е намалена за 6-10 пати, дијаметарот се зголемува на 30 nm.
- ниво на јамка
Нивото на јамката е обезбедено од не-хистонски специфични врзувачки протеини за ДНК кои препознаваат и се врзуваат за специфични секвенци на ДНК, формирајќи јамки од приближно 30-300 kb. Јамката обезбедува генска експресија, т.е. јамката не е само структурна, туку и функционална формација. Скратувањето на ова ниво се јавува 20-30 пати. Дијаметарот се зголемува до 300 nm. Во цитолошките препарати може да се забележат структури во облик на јамка, како што се „четки за ламби“ кај ооцитите од водоземци. Се чини дека овие јамки се превиткани и претставуваат ДНК домени, веројатно што одговараат на единиците на транскрипција и репликација на хроматин. Специфичните протеини ги фиксираат основите на јамките и, можеби, некои од нивните внатрешни делови. Организацијата на доменот слична на јамка промовира преклопување на хроматин во метафазните хромозоми во спирални структури од повисоки редови.
- ниво на домен
Нивото на доменот на организацијата на хромозомот не е доволно проучено. На ова ниво е забележано формирање на домени на јамка - структури на нишки (фибрили) со дебелина од 25-30 nm, кои содржат 60% протеини, 35% ДНК и 5% РНК, практично се невидливи во сите фази од клеточниот циклус со исклучок на митоза и се малку случајно распоредени низ клеточното јадро. Во цитолошките препарати може да се забележат структури во облик на јамка, како што се „четки за ламби“ кај ооцитите од водоземци.
Домените на јамката се прикачени во нивната основа за интрануклеарната протеинска матрица во таканаречените вградени места за прицврстување, честопати наречени MAR/SAR секвенци (MAR, од регионот поврзан со англиската матрица; SAR, од регионите за прицврстување на англиски скеле) - ДНК фрагменти неколку стотици базни парови во должина кои се карактеризираат со висока содржина (>65%) на A/T нуклеотидни парови. Се чини дека секој домен има единствено потекло на репликација и функционира како автономна суперспирална единица. Секој домен на јамка содржи многу единици за транскрипција, чие функционирање е веројатно координирано - целиот домен е или во активна или неактивна состојба.
На ниво на домен, како резултат на секвенцијално хроматинско пакување, се јавува намалување на линеарните димензии на ДНК за приближно 200 пати (700 nm).
- хромозомско ниво
На хромозомско ниво, кондензацијата на профазниот хромозом во метафазен хромозом се случува со набивање на домени на јамката околу аксијалната рамка на нехистонските протеини. Овој суперсвиткување е придружен со фосфорилација на сите H1 молекули во клетката. Како резултат на тоа, метафазниот хромозом може да се прикаже како густо набиени соленоидни јамки, намотани во цврста спирала. Типичен човечки хромозом може да содржи до 2.600 јамки. Дебелината на таквата структура достигнува 1400 nm (две хроматиди), а молекулата на ДНК е скратена за 104 пати, т.е. од 5 cm се протегала ДНК до 5 µm.
Функции на хромозомите
Во интеракција со екстрахромозомските механизми, хромозомите обезбедуваат
- складирање на наследни информации
- користејќи ги овие информации за создавање и одржување на мобилната организација
- регулирање на читањето на наследни информации
- самодуплирање на генетскиот материјал
- трансфер на генетски материјал од матичната клетка во клетките ќерки.
Постојат докази дека кога се активира регион на хроматин, т.е. за време на транскрипцијата, прво хистон H1, а потоа хистонскиот октет се реверзибилно отстранети од него. Ова предизвикува декондензација на хроматин, секвенцијална транзиција на хроматински фибрил од 30 nm во фибрил од 10 nm и негово понатамошно расплетување во делови од слободната ДНК, т.е. губење на структурата на нуклеозомите.
По откривањето на принципот на молекуларна организација на таква супстанција како ДНК во 1953 година, молекуларната биологија почна да се развива. Понатаму во процесот на истражување, научниците открија како се рекомбинира ДНК, нејзиниот состав и како е структуриран нашиот човечки геном.
Секој ден се случуваат сложени процеси на молекуларно ниво. Како е структурирана молекулата на ДНК, од што се состои? А каква улога играат молекулите на ДНК во клетката? Ајде да разговараме подетално за сите процеси што се случуваат во двојниот синџир.
Што е наследна информација?
Значи, каде сето тоа започна? Во далечната 1868 година го пронашле во јадрата на бактериите. И во 1928 година, Н. Колцов ја изнесе теоријата дека сите генетски информации за жив организам се шифрирани во ДНК. Тогаш Џ.
Што е сепак ДНК? Оваа супстанца се состои од 2 обединети нишки, поточно спирали. Дел од таков синџир со одредени информации се нарекува ген.

ДНК ги складира сите информации за тоа каков вид на протеини ќе се формираат и по кој редослед. ДНК макромолекулата е материјален носител на неверојатно обемни информации, кои се запишуваат во строга низа од поединечни тули - нуклеотиди. Има вкупно 4 нуклеотиди; тие се надополнуваат хемиски и геометриски. Овој принцип на надополнување, или комплементарност, во науката ќе биде опишан подоцна. Ова правило игра клучна улога во кодирањето и декодирањето на генетските информации.
Бидејќи нишката на ДНК е неверојатно долга, нема повторувања во оваа низа. Секое живо суштество има своја уникатна нишка на ДНК.
Функции на ДНК
Функциите вклучуваат складирање на наследни информации и нивно пренесување на потомството. Без оваа функција, геномот на еден вид не би можел да се зачува и развие во текот на илјадници години. Организмите кои претрпеле тешки генски мутации имаат поголема веројатност да не преживеат или да ја изгубат способноста да создаваат потомство. Така настанува природна заштита од дегенерација на видот.

Друга суштинска функција е имплементацијата на зачуваните информации. Клетката не може да создаде ниту еден витален протеин без оние инструкции кои се складирани во двоен синџир.
Состав на нуклеинска киселина
Сега со сигурност е познато од што се направени самите нуклеотиди - градежните блокови на ДНК. Тие содржат 3 супстанции:
- Ортофосфорна киселина.
- Азотна основа. Пиримидински бази - кои имаат само еден прстен. Тие вклучуваат тимин и цитозин. Пурински бази, кои содржат 2 прстени. Тоа се гванин и аденин.
- Сахароза. ДНК содржи деоксирибоза, РНК содржи рибоза.
Бројот на нуклеотиди е секогаш еднаков на бројот на азотни бази. Во посебни лаборатории, нуклеотидот се разложува и од него се изолира азотна база. Така се проучуваат индивидуалните својства на овие нуклеотиди и можните мутации во нив.
Нивоа на организација на наследни информации
Постојат 3 нивоа на организација: генетско, хромозомско и геномско. Сите информации потребни за синтеза на нов протеин се содржани во мал дел од синџирот - генот. Односно, генот се смета за најниско и наједноставно ниво на кодирање на информации.

Гените, пак, се составуваат во хромозоми. Благодарение на оваа организација на носителот на наследниот материјал, групите на карактеристики се менуваат според одредени закони и се пренесуваат од една генерација на друга. Треба да се напомене дека во телото има неверојатен број гени, но информациите не се губат дури и кога се рекомбинираат многу пати.
Постојат неколку видови на гени:
- Според нивната функционална намена, постојат 2 вида: структурни и регулаторни секвенци;
- Врз основа на нивното влијание врз процесите што се случуваат во клетката, тие разликуваат: супервитални, смртоносни, условно смртоносни гени, како и мутаторни и антимутаторни гени.
Гените се распоредени долж хромозомот во линеарен редослед. Во хромозомите, информациите не се фокусирани случајно, постои одреден редослед. Постои дури и мапа која ги прикажува позициите или местата на гените. На пример, познато е дека хромозомот бр. 18 шифрира податоци за бојата на очите на детето.
Што е геном? Ова е името дадено на целиот сет на нуклеотидни секвенци во клетката на организмот. Геномот карактеризира цел вид, а не поединец.
Кој е човечкиот генетски код?
Факт е дека целиот огромен потенцијал на човековиот развој лежи веќе во периодот на зачнувањето. Сите наследни информации кои се неопходни за развој на зиготот и растот на детето по раѓањето се шифрирани во гените. ДНК-секциите се најосновните носители на наследни информации.
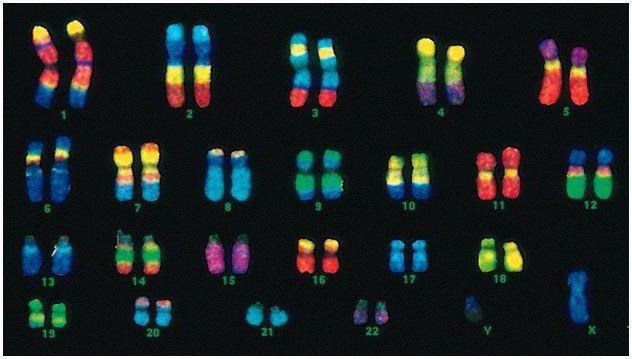
Луѓето имаат 46 хромозоми или 22 соматски парови плус по еден хромозом што го одредува полот од секој родител. Овој диплоиден сет на хромозоми го шифрира целиот физички изглед на една личност, неговите ментални и физички способности и подложност на болести. Соматските хромозоми однадвор не се разликуваат, но носат различни информации, бидејќи едниот од нив е од таткото, другиот од мајката.
Машкиот код се разликува од женскиот код во последниот пар на хромозоми - XY. Женскиот диплоиден сет е последниот пар, XX. Мажјаците добиваат еден Х хромозом од нивната биолошка мајка, кој потоа им се пренесува на нивните ќерки. Половиот хромозом Y се пренесува на синовите.
Човечките хромозоми се многу различни по големина. На пример, најмалиот пар на хромозоми е број 17. И најголемиот пар е 1 и 3.
Дијаметарот на двојната спирала кај луѓето е само 2 nm. ДНК е толку цврсто намотана што се вклопува во малото јадро на клетката, иако би била долга до 2 метри доколку не се извитка. Должината на спиралата е стотици милиони нуклеотиди.
Како се пренесува генетскиот код?
Значи, каква улога играат молекулите на ДНК во клеточната делба? Гените - носители на наследни информации - се наоѓаат во секоја клетка од телото. За да го пренесат својот код на организам ќерка, многу суштества ја делат својата ДНК на 2 идентични спирали. Ова се нарекува репликација. За време на процесот на репликација, ДНК се одмотува и специјални „машини“ ја завршуваат секоја нишка. Откако генетската спирала ќе се раздели, јадрото и сите органели почнуваат да се делат, а потоа и целата клетка.
Но, луѓето имаат поинаков процес на пренос на ген - сексуален. Карактеристиките на таткото и мајката се измешани, новиот генетски код содржи информации од двајцата родители.
Складирањето и преносот на наследни информации е можно поради сложената организација на спиралата на ДНК. На крајот на краиштата, како што рековме, структурата на протеините е шифрирана во гените. Откако ќе се создаде во моментот на зачнувањето, овој код ќе се копира во текот на животот. Кариотипот (личен сет на хромозоми) не се менува за време на обновувањето на клетките на органите. Преносот на информации се врши со помош на полови гамети - машки и женски.
Само вирусите кои содржат една нишка на РНК не се способни да ги пренесат своите информации до нивните потомци. Затоа, за репродукција им се потребни човечки или животински клетки.
Имплементација на наследни информации
Во клеточното јадро постојано се случуваат важни процеси. Сите информации запишани во хромозомите се користат за изградба на протеини од амино киселини. Но, синџирот на ДНК никогаш не го напушта јадрото, па затоа му е потребна помош од друго важно соединение: РНК. РНК е способна да навлезе во нуклеарната мембрана и да комуницира со синџирот на ДНК.
Преку интеракцијата на ДНК и 3 типа на РНК се реализираат сите кодирани информации. На кое ниво се случува имплементацијата на наследни информации? Сите интеракции се случуваат на ниво на нуклеотиди. Месинџер РНК копира дел од синџирот на ДНК и ја носи оваа копија до рибозомот. Тука започнува синтезата на нова молекула од нуклеотиди.
За да може mRNA да го копира потребниот дел од синџирот, спиралата се расплетува и потоа, по завршувањето на процесот на прекодирање, повторно се обновува. Покрај тоа, овој процес може да се случи истовремено на 2 страни од 1 хромозом.
Принцип на комплементарност
Тие се состојат од 4 нуклеотиди - аденин (А), гванин (G), цитозин (Ц), тимин (Т). Тие се поврзани со водородни врски според правилото за комплементарност. Работата на Е. Чаргаф помогна да се воспостави ова правило, бидејќи научникот забележал некои обрасци во однесувањето на овие супстанции. Е. Чаргаф открил дека моларниот однос на аденин и тимин е еднаков на еден. И на ист начин, односот на гванин и цитозин е секогаш еднаков на еден.
Врз основа на неговата работа, генетичарите формираа правило за интеракцијата на нуклеотидите. Правилото за комплементарност вели дека аденин се комбинира само со тимин, а гванин само со цитозин. За време на декодирањето на спиралата и синтезата на нов протеин во рибозомот, ова правило на алтернација помага брзо да се најде потребната амино киселина која е прикачена на трансферната РНК.
РНК и нејзините типови
Што е наследна информација? нуклеотиди во двојна нишка на ДНК. Што е РНК? Која е нејзината работа? РНК, или рибонуклеинска киселина, помага да се извлечат информации од ДНК, да се декодираат и, врз основа на принципот на комплементарност, да се создадат протеини неопходни за клетките.
Вкупно има 3 типа на РНК. Секој од нив врши строго своја функција.
- Информативни (mRNA), или уште наречена матрица. Тој оди директно во центарот на клетката, во јадрото. Во еден од хромозомите го наоѓа потребниот генетски материјал за да изгради протеин и копира една од страните на двојната жичка. Копирањето се случува повторно според принципот на комплементарност.
- Транспорте мала молекула која има нуклеотидни декодери на едната страна, а аминокиселини што одговараат на основниот код на другата страна. Задачата на tRNA е да ја достави до „работилницата“, односно до рибозомот, каде што ја синтетизира потребната амино киселина.
- rRNA е рибозомална.Ја контролира количината на протеин што се произведува. Се состои од 2 дела - аминокиселина и пептиден дел.
Единствената разлика во декодирањето е тоа што РНК нема тимин. Наместо тимин, тука е присутен урацил. Но, тогаш, за време на процесот на синтеза на протеини, tRNA сè уште правилно ги инсталира сите амино киселини. Ако се појават какви било неуспеси во декодирањето на информациите, тогаш се јавува мутација.
Поправка на оштетена молекула на ДНК
Процесот на обновување на оштетената двојна жичка се нарекува поправка. За време на процесот на поправка, оштетените гени се отстрануваат.

Потоа, потребната низа на елементи точно се репродуцира и се сече на истото место на синџирот од каде што е отстранет. Сето ова се случува благодарение на специјалните хемикалии - ензими.
Зошто се јавуваат мутации?
Зошто некои гени почнуваат да мутираат и престануваат да ја вршат својата функција - складирање на витални наследни информации? Ова се случува поради грешка во декодирањето. На пример, ако аденин случајно се замени со тимин.
Постојат и хромозомски и геномски мутации. Хромозомските мутации се случуваат кога делови од наследни информации се губат, се дуплираат или дури се префрлаат и се вметнуваат во друг хромозом.

Геномските мутации се најсериозни. Нивната причина е промена на бројот на хромозоми. Односно, кога наместо пар - диплоидно множество, во кариотипот е присутен триплоиден сет.
Најпознатиот пример за триплоидна мутација е Даунов синдром, кај кој личниот сет на хромозоми е 47. Кај таквите деца на местото на 21-от пар се формираат 3 хромозоми.
Исто така, постои позната мутација наречена полиплоидија. Но, полиплоидијата се јавува само кај растенијата.
Молекулата на ДНК се состои од две нишки кои формираат двојна спирала. Нејзината структура првпат беше дешифрирана од Френсис Крик и Џејмс Вотсон во 1953 година.
Отпрвин, молекулата на ДНК, која се состои од пар нуклеотидни синџири извиткани еден околу друг, предизвика прашања за тоа зошто ја има оваа конкретна форма. Научниците овој феномен го нарекуваат комплементарност, што значи дека само одредени нуклеотиди можат да се најдат еден спроти друг во неговите нишки. На пример, аденинот е секогаш спротивен тимин, а гванин е секогаш спротивен на цитозин. Овие нуклеотиди на молекулата на ДНК се нарекуваат комплементарни.
Шематски тоа е прикажано вака:
Т - А
Ц - Г
Овие парови формираат хемиска нуклеотидна врска, која го одредува редоследот на амино киселините. Во првиот случај е малку послаб. Врската помеѓу C и G е посилна. Некомплементарните нуклеотиди не формираат парови едни со други.
За зградата
Значи, структурата на молекулата на ДНК е посебна. Тој ја има оваа форма со причина: факт е дека бројот на нуклеотиди е многу голем, а потребен е многу простор за да се сместат долгите синџири. Тоа е поради оваа причина што синџирите се карактеризираат со спирален пресврт. Овој феномен се нарекува спирализација, овозможува нишките да се скратат за околу пет до шест пати.
Телото користи некои молекули од овој тип многу активно, други ретко. Вторите, покрај спирализацијата, исто така се подложени на такво „компактно пакување“ како суперспирализација. И тогаш должината на молекулата на ДНК се намалува за 25-30 пати.

Какво е „пакувањето“ на молекулата?
Процесот на превиткување вклучува хистонски протеини. Имаат структура и изглед на калем од конец или прачка. На нив се намотани спирализирани нишки, кои веднаш стануваат „компактно спакувани“ и заземаат малку простор. Кога ќе се појави потреба да се користи една или друга нишка, таа се одмотува од калем, на пример, хистонски протеин, а спиралата се одмотува во два паралелни синџири. Кога молекулата на ДНК е во оваа состојба, од неа може да се прочитаат потребните генетски податоци. Сепак, постои еден услов. Добивањето информации е можно само ако структурата на молекулата на ДНК има неизвртена форма. Хромозомите кои се достапни за читање се нарекуваат еухроматини, а ако се супернамотани, тогаш тие се веќе хетерохроматини.
Нуклеински киселини
Нуклеинските киселини, како и протеините, се биополимери. Главната функција е складирање, имплементација и пренос на наследни (генетски информации). Тие доаѓаат во два вида: ДНК и РНК (деоксирибонуклеинска и рибонуклеинска). Мономерите во нив се нуклеотиди, од кои секој содржи остаток на фосфорна киселина, шеќер од пет јаглерод (деоксирибоза/рибоза) и азотна база. Кодот на ДНК вклучува 4 типа нуклеотиди - аденин (А) / гванин (G) / цитозин (C) / тимин (Т). Тие се разликуваат по азотната база што ја содржат.
Во молекулата на ДНК, бројот на нуклеотиди може да биде огромен - од неколку илјади до десетици и стотици милиони. Ваквите џиновски молекули можат да се испитаат преку електронски микроскоп. Во овој случај, ќе можете да видите двоен синџир на полинуклеотидни нишки, кои се поврзани едни со други со водородни врски на азотни бази на нуклеотидите.

Истражување
Во текот на истражувањето, научниците открија дека типовите на молекули на ДНК се разликуваат кај различни живи организми. Исто така, беше откриено дека гванин од еден синџир може да се врзе само за цитозин, а тимин за аденин. Распоредот на нуклеотидите во еден синџир строго одговара на паралелниот. Благодарение на оваа комплементарност на полинуклеотидите, молекулата на ДНК е способна за удвојување и саморепродукција. Но, прво, комплементарните синџири, под влијание на специјални ензими кои ги уништуваат спарените нуклеотиди, се разминуваат, а потоа во секој од нив започнува синтезата на исчезнатиот синџир. Ова се случува поради слободните нуклеотиди присутни во големи количини во секоја клетка. Како резултат на ова, наместо „мајчината молекула“, се формираат две „ќерки“, идентични по состав и структура, а кодот на ДНК станува оригинален. Овој процес е претходник на клеточната делба. Обезбедува пренос на сите наследни податоци од матичните клетки до ќерките, како и до сите наредни генерации.
Како се чита генскиот код?
Денес, не се пресметува само масата на молекулата на ДНК - можно е да се дознаат и посложени податоци кои претходно биле недостапни за научниците. На пример, можете да прочитате информации за тоа како еден организам ја користи сопствената клетка. Се разбира, на почетокот оваа информација е во шифрирана форма и има форма на одредена матрица и затоа мора да се транспортира до посебен носач, а тоа е РНК. Рибонуклеинската киселина може да навлезе во клетката преку нуклеарната мембрана и да ги чита кодираните информации внатре. Така, РНК е носител на скриени податоци од јадрото до клетката и се разликува од ДНК по тоа што содржи рибоза наместо деоксирибоза и урацил наместо тимин. Покрај тоа, РНК е едноверижна.

Синтеза на РНК
Длабинската анализа на ДНК покажа дека откако РНК ќе го напушти јадрото, таа влегува во цитоплазмата, каде што може да се интегрира како матрица во рибозомите (специјални ензимски системи). Водени од добиените информации, тие можат да синтетизираат соодветна низа на протеински амино киселини. Рибозомот учи од тројниот код кој тип на органско соединение треба да се прикачи на синџирот на протеини. Секоја аминокиселина има своја специфична тројка, која ја шифрира.
Откако ќе заврши формирањето на синџирот, тој добива специфична просторна форма и се претвора во протеин способен да ги извршува своите хормонски, градежни, ензимски и други функции. За секој организам тоа е генски производ. Од него се одредуваат сите видови квалитети, својства и манифестации на гените.

Гените
Процесите на секвенционирање првенствено беа развиени за да се добијат информации за тоа колку гени има една молекула на ДНК во својата структура. И, иако истражувањата им овозможија на научниците да постигнат голем напредок во ова прашање, сè уште не е можно да се знае нивниот точен број.
Пред само неколку години се претпоставуваше дека молекулите на ДНК содржат приближно 100 илјади гени. Малку подоцна, бројката се намали на 80 илјади, а во 1998 година, генетичарите изјавија дека во една ДНК се присутни само 50 илјади гени, што е само 3% од вкупната должина на ДНК. Но, најновите заклучоци на генетичарите беа впечатливи. Сега тие тврдат дека геномот вклучува 25-40 илјади од овие единици. Излегува дека само 1,5% од хромозомската ДНК е одговорна за кодирање на протеините.
Истражувањето не застана тука. Паралелен тим од специјалисти за генетски инженеринг открил дека бројот на гени во една молекула е точно 32 илјади. Како што можете да видите, сè уште е невозможно да се добие дефинитивен одговор. Има премногу противречности. Сите истражувачи се потпираат само на нивните резултати.
Имаше ли еволуција?
И покрај фактот дека нема докази за еволуцијата на молекулата (бидејќи структурата на молекулата на ДНК е кревка и мала по големина), научниците сепак направија една претпоставка. Врз основа на лабораториски податоци, тие ја изразија следнава верзија: во почетната фаза на нејзиниот изглед, молекулата имаше форма на едноставен пептид што се реплицира, кој вклучуваше до 32 амино киселини пронајдени во античките океани.
По саморепликацијата, благодарение на силите на природната селекција, молекулите се здобија со способност да се заштитат од надворешни елементи. Почнаа да живеат подолго и да се размножуваат во поголеми количини. Молекулите кои се најдоа во липидниот меур ги имаа сите шанси да се репродуцираат. Како резултат на низа последователни циклуси, липидните меурчиња добија форма на клеточни мембрани, а потоа - добро познатите честички. Треба да се забележи дека денес секој дел од молекулата на ДНК е сложена и јасно функционална структура, чиишто карактеристики научниците сè уште не ги проучиле целосно.

Модерен свет
Неодамна, научници од Израел развија компјутер кој може да врши трилиони операции во секунда. Денес тоа е најбрзиот автомобил на Земјата. Целата тајна е дека иновативниот уред се напојува со ДНК. Професорите велат дека во блиска иднина ваквите компјутери дури ќе можат да генерираат енергија.
Пред една година, специјалисти од Институтот Вајцман во Реховот (Израел) најавија создавање на програмабилна молекуларна компјутерска машина која се состои од молекули и ензими. Тие ги замениле силиконските микрочипови со нив. До денес, тимот има понатамошен напредок. Сега само една молекула на ДНК може да му обезбеди на компјутерот потребните податоци и потребното гориво.
Биохемиските „нанокомпјутери“ не се фикција, тие веќе постојат во природата и се манифестираат во секое живо суштество. Но, често тие не се управувани од луѓе. Едно лице сè уште не може да работи на геномот на кое било растение за да го пресмета, да речеме, бројот „Пи“.
Идејата за користење на ДНК за складирање/обработка на податоци првпат им падна на ум на научниците во 1994 година. Тогаш беше искористена молекула за решавање на едноставен математички проблем. Оттогаш, голем број истражувачки групи предложија различни проекти поврзани со ДНК компјутерите. Но, овде сите обиди беа засновани само на енергетската молекула. Не можете да видите таков компјутер со голо око, тој изгледа како проѕирен раствор од вода во епрувета. Во него нема механички делови, туку само трилиони биомолекуларни уреди - и ова е само во една капка течност!
Човечка ДНК
Луѓето станаа свесни за типот на човечка ДНК во 1953 година, кога научниците за прв пат успеаја да му покажат на светот двоверижен модел на ДНК. За ова, Кирк и Вотсон ја добија Нобеловата награда, бидејќи ова откритие стана фундаментално во 20 век.
Со текот на времето, се разбира, тие докажаа дека структурираната човечка молекула може да изгледа не само како во предложената верзија. Откако направија подетална ДНК анализа, тие ја открија формата А-, Б- и левата рака З-. Формата А- често е исклучок, бидејќи се формира само ако има недостаток на влага. Но, тоа е можно само во лабораториски студии; за природната средина ова е аномално; таков процес не може да се случи во жива клетка.
Обликот Б е класичен и е познат како синџир со двојна десна рака, но формата Z не само што е извиткана во спротивна насока од лево, туку има и повеќе цик-цак изглед. Научниците, исто така, ја идентификуваа формата G-quadruplex. Неговата структура нема 2, туку 4 нишки. Според генетичарите, оваа форма се јавува во области каде што има вишок на гванин.

Вештачка ДНК
Денес веќе постои вештачка ДНК, која е идентична копија на вистинската; совршено ја следи структурата на природната двојна спирала. Но, за разлика од оригиналниот полинуклеотид, вештачкиот има само два дополнителни нуклеотиди.
Бидејќи синхронизацијата е создадена врз основа на информации добиени од различни студии за вистинска ДНК, може да се копира, да се самореплицира и да се развива. Експертите работат на создавање на ваква вештачка молекула околу 20 години. Резултатот е неверојатен изум кој може да го користи генетскиот код на ист начин како и природната ДНК.
На четирите постоечки азотни бази, генетичарите додадоа две дополнителни, кои беа создадени со хемиска модификација на природните бази. За разлика од природната ДНК, вештачката ДНК се покажа како прилично кратка. Содржи само 81 базни парови. Сепак, тој исто така се репродуцира и еволуира.
Репликацијата на вештачки добиена молекула се одвива благодарение на полимеразната верижна реакција, но засега тоа не се случува независно, туку со интервенција на научниците. Тие самостојно ги додаваат потребните ензими на споменатата ДНК, ставајќи ја во специјално подготвен течен медиум.
Конечниот резултат
Процесот и конечниот исход на развојот на ДНК може да бидат под влијание на различни фактори, како што се мутациите. Ова го прави неопходно да се проучуваат примероци од материја, така што резултатот од анализата е сигурен и сигурен. Пример е тест за татковство. Но, не можеме а да не се радуваме што инцидентите како што е мутацијата се ретки. Сепак, примероците од материјата секогаш се проверуваат со цел да се добијат попрецизни информации врз основа на анализата.

Растителна ДНК
Благодарение на технологиите за високо секвенционирање (HTS), направена е револуција во областа на геномијата - можно е и екстракција на ДНК од растенија. Се разбира, добивањето висококвалитетна молекуларна ДНК од растителен материјал предизвикува одредени потешкотии поради големиот број копии на ДНК на митохондриите и хлоропластите, како и високото ниво на полисахариди и фенолни соединенија. За да се изолира структурата што ја разгледуваме во овој случај, се користат различни методи.
Водородна врска во ДНК
Водородната врска во молекулата на ДНК е одговорна за електромагнетната привлечност создадена помеѓу позитивно наелектризираниот водороден атом кој е прикачен на електронегативен атом. Оваа диполна интеракција не го исполнува критериумот за хемиска врска. Но, тоа може да се појави интермолекуларно или во различни делови на молекулата, т.е. интрамолекуларно.

Атом на водород се прикачува на електронегативниот атом кој е донатор на врската. Електронегативен атом може да биде азот, флуор или кислород. Тој - преку децентрализација - го привлекува електронскиот облак од водородното јадро кон себе и го прави водородниот атом (делумно) позитивно наелектризиран. Бидејќи големината на H е мала во споредба со другите молекули и атоми, полнењето е исто така мало.
Декодирање на ДНК
Пред да дешифрираат молекула на ДНК, научниците прво земаат огромен број клетки. За најточна и најуспешна работа потребни се околу милион од нив. Резултатите добиени во текот на студијата постојано се споредуваат и евидентираат. Денес, декодирањето на геномот повеќе не е реткост, туку пристапна процедура.

Се разбира, дешифрирањето на геномот на една клетка е непрактична вежба. Податоците добиени за време на ваквите студии не се од интерес за научниците. Но, важно е да се разбере дека сите тековно постоечки методи за декодирање, и покрај нивната сложеност, не се доволно ефикасни. Тие ќе дозволат само читање на 40-70% од ДНК.
Сепак, професорите од Харвард неодамна најавија метод преку кој може да се дешифрира 90% од геномот. Техниката се заснова на додавање прајмерски молекули во изолираните клетки, со чија помош започнува репликацијата на ДНК. Но, дури и овој метод не може да се смета за успешен; сепак треба да се рафинира пред да може отворено да се користи во науката.
