DNA-molekylet er lokalisert. DNA og gener. Nytt blikk på DNA
Biokjemiske arvegrunnlag.
Nukleinsyrers genetiske rolle.
Nukleinsyrer er biologiske polymerer som finnes i alle celler, fra primitive til komplekse. De ble først oppdaget av Johann Friedrich Miescher i 1868 i celler rike på kjernefysisk materiale (leukocytter, laksesæd). Begrepet "nukleinsyrer" ble foreslått i 1889.
Det finnes to typer nukleinsyrer: DNA, RNA (ATP - mononukleotid). DNA og RNA er molekyler - matriser. DNA er inneholdt ca. 6 * 10 -12 g i somatiske celler: i kjernen, mitokondrier. RNA er en del av ribosomer og finnes i kjernen og cytoplasma.
Studien og beviset på den ledende rollen til nukleinsyrer i overføringen av arvelig informasjon ble utført på virale partikler. Tobakksmosaikkvirus er kjent for å være virulent for tobakk og groblad. Den virale partikkelen består av 95 % protein og 5 % nukleinsyre. Proteinkapsidet i viruspartiklene ble byttet ut, men etter en tid ble proteinet i begge stammer transformert til sin tidligere form.
I bakteriofager som infiserer E. coli ble fagkappeproteinene merket med radioaktivt S, og fag-DNA ble merket med radioaktivt P. I en bakteriecelle infisert med fagen ble det dannet fagpartikler som kun inneholdt radioaktivt P.
Struktur og funksjoner til DNA- og RNA-molekyler.
Nukleinsyrer er biopolymerer med uregelmessig struktur, hvis monomerer er nukleotider. Nukleotidet består av rester av tre stoffer: fosforsyre, karbohydrat - pentose, nitrogenholdig base. DNA-nukleotider inneholder karbohydratet deoksyribose, og RNA inneholder ribose. Rester av purin og pyrimidin nitrogenholdige baser som utgjør DNA er adenin, guanin, cytosin, tymin. RNA-molekyler inneholder adenin, guanin, cytosin og uracil.
Nukleotider er koblet til hverandre gjennom fosforsyreresten til ett nukleotid og karbohydratet til et annet ved en sterk kovalent esterbinding, kalt en "oksygenbro". Bindingen går gjennom det 5. karbonatomet i et karbohydrat i ett nukleotid til det tredje karbonatomet i et karbohydrat i et annet nukleotid. Nukleotidsekvensen representerer den primære strukturen til nukleinsyrer. RNA er en enkelt polynukleotidkjede. DNAs struktur er en dobbel polynukleotidkjede, kveilet inn i en spiral.
Den sekundære strukturen til DNA dannes når en andre DNA-streng dukker opp, arrangert i henhold til prinsippet om komplementaritet med hensyn til den første. Den andre kjeden er motsatt av den første (antiparallell). De nitrogenholdige basene ligger i et plan vinkelrett på molekylets plan - dette ligner en spiraltrapp. Rekkverket til denne trappen er fosforsyrerester og karbohydrater, og trinnene er nitrogenholdige baser.
De nitrogenholdige basene som utgjør hvert nukleotid i motsatt rettede kjeder er i stand til å danne komplementære hydrogenbindinger med hverandre (på grunn av de eksisterende funksjonelle gruppene i strukturen til hver nitrogenholdig base). Adenylnukleotid er komplementært til tymin, guanyl til cytosin og omvendt. I seg selv er disse bindingene skjøre, men et DNA-molekyl "sydd" gjentatte ganger langs hele lengden med slike bindinger representerer en veldig sterk forbindelse.
Komplementaritet- dette er den romlig-strukturelle og kjemiske korrespondansen mellom nitrogenholdige baser til hverandre; de passer til hverandre "som en nøkkel til en lås."
Ett DNA-molekyl kan inneholde 108 eller flere nukleotider.
Strukturen til DNA-molekylet som en dobbel antiparallell helix ble foreslått i 1953 av den amerikanske biologen James Watson og den engelske fysikeren Francis Crick.
DNA-molekylet til enhver levende organisme på planeten består av bare fire typer nukleotider, som er forskjellige fra hverandre i de nitrogenholdige basene de inneholder: adenyl, guanyl, tymin og cytosin. I det allsidighet DNA. Rekkefølgen deres er forskjellig, og antallet er uendelig.
For hver art av levende organismer og for hver organisme separat, er denne sekvensen individuell og strengt tatt spesifikk .
Egenhet Strukturen til DNA er at de kjemisk aktive delene av molekylet - nitrogenholdige baser, er nedsenket i midten av helixen og danner komplementære bindinger med hverandre, og deoksyribose- og fosforsyrerester er lokalisert i periferien og dekker tilgang til nitrogenholdige baser - de er kjemisk inaktive. Denne strukturen kan opprettholde kjemisk stabilitet i lang tid. Hva mer er nødvendig for å lagre arvelig informasjon? Det er disse egenskapene til DNA-strukturen som bestemmer dens evne til å kode og reprodusere genetisk informasjon.
Den sterke strukturen til DNA er ganske vanskelig å ødelegge. Likevel skjer dette regelmessig i cellen – under syntesen av RNA og doblingen av selve DNA-molekylet før celledeling.
Duplisering, DNA-replikasjon begynner med det faktum at et spesielt enzym - DNA-polymerase - vikler ut den doble helixen og skiller den i separate tråder - det dannes en reduplikasjonsgaffel. Enzymet fungerer som en lås på en glidelås. Ved hver enkelttrådet kjede - de klebrige endene av reduplikasjonsgaffelen - syntetiseres en ny kjede fra frie nukleotider i karyoplasma i henhold til komplementaritetsprinsippet. I to nye DNA-molekyler forblir en streng den opprinnelige moderstrengen, og den andre forblir den nye datterstrengen. Som et resultat, i stedet for ett DNA-molekyl, vises to molekyler med samme eksakte nukleotidsammensetning som det opprinnelige.
I levende systemer møter vi en ny type reaksjoner ukjent i den livløse naturen. De heter matrisesyntesereaksjoner . Matrisesyntese er som å støpe på en matrise: nye molekyler syntetiseres i nøyaktig samsvar med planen som er lagt i strukturen til eksisterende molekyler. Disse reaksjonene sikrer den nøyaktige sekvensen av monomerenheter i de syntetiserte polymerene. Monomerer kommer til et bestemt sted på molekyler som fungerer som en matrise der reaksjonen finner sted. Hvis slike reaksjoner skjedde som et resultat av tilfeldige kollisjoner av molekyler, ville de forløpe uendelig sakte. Syntesen av komplekse molekyler basert på malprinsippet utføres raskt og nøyaktig ved hjelp av enzymer. Templatesyntese ligger til grunn for de viktigste reaksjonene i syntesen av nukleinsyrer og proteiner. Rollen til matrisen i cellen spilles av molekyler av nukleinsyrer DNA eller RNA. Monomere molekyler som polymeren er syntetisert fra - nukleotider eller aminosyrer - i samsvar med komplementaritetsprinsippet, er lokalisert og fiksert på matrisen i en strengt definert rekkefølge. Deretter kobles monomerenhetene til en polymerkjede, og den ferdige polymeren forlater matrisen. Etter dette er matrisen klar for montering av et nytt, nøyaktig det samme polymermolekylet.
Matrise-type reaksjoner er et spesifikt trekk ved en levende celle. De er grunnlaget for den grunnleggende egenskapen til alle levende ting - evnen til å reprodusere sin egen type.
Funksjoner av nukleinsyrer– lagring og overføring av arvelig informasjon. DNA-molekyler koder for informasjon om proteinets primære struktur. Syntesen av mRNA-molekyler foregår på DNA-matrisen. Denne prosessen kalles "transkripsjon". I-RNA i prosessen med "oversettelse" implementerer informasjon i form av en sekvens av aminosyrer i et proteinmolekyl.
DNAet til hver celle bærer ikke bare informasjon om de strukturelle proteinene som bestemmer cellens form, men også om alle enzymproteiner, hormonproteiner og andre proteiner, samt strukturen til alle typer RNA.
Kanskje nukleinsyrer gir ulike typer biologisk minne - immunologisk, nevrologisk, etc., og spiller også en betydelig rolle i reguleringen av biosyntetiske prosesser.
Relatert informasjon.
Det første beviset på rollen til DNA som bærer av arvelig informasjon i organismer vakte enorm oppmerksomhet til studiet av nukleinsyrer. I 1869 isolerte F. Miescher et spesielt stoff fra cellekjerner, som han kalte nuklein. Etter 20 år ble dette navnet erstattet av begrepet nukleinsyre. I 1924 utviklet R. Felgen en metode for cytologisk gjenkjennelse av nukleinsyrer gjennom deres spesifikke farging og viste at DNA er lokalisert i cellekjerner, og RNA i cytoplasma. I 1936 A.N. Belozersky og I.I. Dubrovskaya isolerte DNA i sin rene form fra kjernene til planteceller. På begynnelsen av 1930-tallet. De grunnleggende kjemiske prinsippene for strukturen til nukleinsyresukker ble belyst, og i 1953 ble det laget en strukturell modell av DNA.
Den grunnleggende strukturelle enheten til nukleinsyrer er nukleotid, som består av tre kjemisk forskjellige deler forbundet med kovalente bindinger (fig. 5.2).
Ris. 5.2. Strukturformler: EN- nukleotider; b- DNA; V - RNA (se også s. 110)

Ris. 5.2. Slutt. Strukturformler: EN- nukleotider; 6 - DNA; V- RNA
Den første delen er sukker som inneholder fem karbonatomer: deoksyribose i DNA og ribose i RNA.
Den andre delen av nukleotidet, en nitrogenholdig purin- eller pyrimidinbase, er kovalent knyttet til det første karbonatomet i sukkeret, og danner en struktur som kalles nukleosid. DNA inneholder purinbaser - adenin(A) og guanin(D) - og pyrimidinbaser - tymin(T) og cytosin(C). De tilsvarende nukleosidene kalles deoksyadenosin, deoksyguanosin, deoksytymidin og deoksycytidin. RNA inneholder de samme purinbasene som DNA, en pyrimidinbase cytosin, og i stedet for tymin inneholder den uracil(U); de tilsvarende nukleosidene kalles adenosin, guanosin, uridin og cytidin.
Den tredje delen av nukleotidet er en fosfatgruppe, som forbinder nabonukleosider til en polymerkjede gjennom fosfodiesterbindinger mellom 5-karbonatomet til ett sukker og 3-karbonatomet til et annet (fig. 5.2, b, V). Nukleotider kalles nukleosider med en eller flere fosfatgrupper festet med esterbindinger til sukkerets 3" eller 5 karbonatomer. Syntesen av nukleotider går foran syntesen av nukleinsyrer; følgelig er nukleotider produkter av kjemisk eller enzymatisk hydrolyse av nukleinsyrer.
Nukleinsyrer er svært lange polymerkjeder som består av mononukleotider forbundet med 5- og 3'-fosfodiesterbindinger. Et intakt DNA-molekyl inneholder, avhengig av type organisme, fra flere tusen til mange millioner nukleotider, et intakt RNA-molekyl - fra 100 til 100 tusen eller flere nukleotider.
Resultatene av E. Chargaffs analyser av nukleotidsammensetningen til DNA av ulike artsformer viste at molekylforholdet mellom ulike nitrogenholdige baser – adenin, guanin, tymin, cytosin – varierer mye. Følgelig ble det bevist at DNA slett ikke er en monoton polymer bestående av identiske tetranukleotider, slik man antok på 40-tallet. XX århundrer, og at den fullt ut besitter kompleksiteten som er nødvendig for bevaring og overføring av arvelig informasjon i form av en spesifikk sekvens av nukleotidbaser.
Forskning av E. Chargaff avslørte også en egenskap som er iboende i alle DNA-molekyler: molinnholdet av adenin er lik innholdet av tymin, og molinnholdet av guanin er lik innholdet av cytosin. Disse likhetene kalles Chargaffs ekvivalensregel: [A] = [T], [G] = [C]; antall puriner er lik antall pyrimidiner. Avhengig av art er det kun forholdet ([A] + [T])/([G] + [C]) som endres (tabell 5.1).
|
Sammensetning av baser |
Holdning |
Asymmetri |
|||||
|
grunner |
|||||||
|
(A + T)/(G + C) |
|||||||
|
Dyr |
|||||||
|
Skilpadde |
|||||||
|
sjøkrabbe |
|||||||
|
Kråkebolle |
|||||||
|
Planter, sopp |
|||||||
|
hvetekim |
|||||||
|
Sopp Aspergillus niger |
|||||||
|
Bakterie |
|||||||
|
Escherichia coli |
|||||||
|
Staphylococcus aureus |
|||||||
|
Clostridium perfringens |
|||||||
|
Brucela abort |
|||||||
|
Sarcina lutea |
|||||||
|
Bakteriofager |
|||||||
|
FH 174 (viral form) |
|||||||
|
FH 174 (replikativ form) |
|||||||
Forholdet mellom baser kalles nukleotidkoeffisient(arter) spesifisitet. Chargaffs oppdagelse formulerte et viktig strukturelt trekk ved DNA, som senere ble reflektert i den strukturelle modellen av DNA av J. Watson og F. Crick (1953), som faktisk viste at Chargaffs regler ikke legger noen begrensninger på det mulige antallet kombinasjoner av forskjellige basesekvenser som er i stand til å danne molekyler DNA.
Konseptet med nukleotidspesifisitet dannet grunnlaget for en ny gren av biologi - gensystematikk, som opererer ved å sammenligne sammensetningen og strukturen til nukleinsyrer for å bygge et naturlig system av organismer.
I følge Watson-Crick-modellen består et DNA-molekyl av to polynukleotidkjeder (tråder, tråder), koblet til hverandre ved hjelp av tverrgående hydrogenbindinger mellom nitrogenholdige baser i henhold til komplementærprinsippet (adenin i en kjede er forbundet med to hydrogenbindinger med tymin i den motsatte kjeden, og guanin og cytosin i forskjellige kjeder er forbundet med hverandre med tre hydrogenbindinger). I dette tilfellet er to polynukleotidkjeder av ett molekyl antiparallelle, dvs. motsatt 3"-enden av en kjede er 5"-enden av den andre kjeden og omvendt (fig. 5.3). Imidlertid bør man huske på moderne data om at det genetiske materialet til noen virus er representert av enkelttrådede (enkeltrådede) DNA-molekyler. Basert på røntgendiffraksjonsanalyse av DNA, konkluderte J. Watson og F. Crick også med at det dobbelttrådete molekylet har en sekundær struktur i form av en helix vridd i retning fra venstre til høyre, som senere ble kalt 5. -form (fig. 5.4). Til dags dato er det bevist at det i tillegg til den vanligste 5-formen er mulig å oppdage DNA-seksjoner som har en annen konfigurasjon - som høyrehendte (former EN, C), og vridd fra høyre til venstre (venstre vridd, eller Z-form) (fig. 5.4). Det er visse forskjeller mellom disse formene for sekundær DNA-struktur (tabell 5.2). For eksempel er avstanden mellom to tilstøtende par nitrogenholdige baser i en dobbelttrådet helix, uttrykt i nanometer (nm), preget av forskjellige verdier for 5-formen og Z-formen (henholdsvis 0,34 og 0,38 nm) . I fig. Figur 5.5 viser moderne tredimensjonale modeller av "venstrehendte" og "høyrehendte" former for DNA.

Ris. 5.3. skjematisk representasjon av den primære strukturen til et fragment av et dobbelttrådet DNA-molekyl: A - adenin; G - guanin; T - tymin; C - cytosin

Ris. 5.4.

Tabell 5.2
Egenskaper til ulike former for DNA-dobbeltspiraler
RNA-molekyler, avhengig av deres strukturelle og funksjonelle egenskaper, er delt inn i flere typer: budbringer-RNA (mRNA, eller mRNA), ribosomalt RNA (rRNA), overførings-RNA (tRNA), lite kjernefysisk RNA (snRNA), etc. I motsetning Fra DNA, RNA-molekyler er alltid enkelttrådet (enkeltrådet). Imidlertid kan de danne mer komplekse (sekundære) konfigurasjoner på grunn av den komplementære forbindelsen av individuelle seksjoner av en slik kjede basert på samspillet mellom komplementære nitrogenholdige baser (A-U og G-C). Som et eksempel kan du vurdere kløverbladkonfigurasjonen for fenylalaninoverførings-RNA-molekylet (figur 5.6).

Ris. 5.6.
I 1953 foreslo D. Watson og F. Crick en modell av strukturen til DNA, som var basert på følgende postulater:
- 1. DNA er en polymer som består av nukleotider forbundet med 3" og 5" fosfodiesterbindinger.
- 2. Sammensetningen av DNA-nukleotider følger Chargaffs regler.
- 3. DNA-molekylet har en dobbel helixstruktur, som minner om en spiraltrapp, noe som fremgår av røntgendiffraksjonsmønstre av DNA-tråder først oppnådd av M. Wilkins og R. Franklin.
- 4. Strukturen til polymeren, som vist ved syre-basetitrering av naturlig (naturlig) DNA, stabiliseres av hydrogenbindinger. Titrering og oppvarming av naturlig DNA forårsaker en merkbar endring i dets fysiske egenskaper, spesielt viskositet, og transformerer det til en denaturert form, og kovalente bindinger blir ikke ødelagt.
Nukleinsyrer er høymolekylære stoffer som består av mononukleotider, som er koblet til hverandre i en polymerkjede ved hjelp av 3", 5" fosfodiesterbindinger og er pakket i celler på en bestemt måte.
Nukleinsyrer er biopolymerer av to typer: ribonukleinsyre (RNA) og deoksyribonukleinsyre (DNA). Hver biopolymer består av nukleotider som er forskjellige i karbohydratrestene (ribose, deoksyribose) og en av de nitrogenholdige basene (uracil, tymin). I følge disse forskjellene fikk nukleinsyrer navnet sitt.
Struktur av deoksyribonukleinsyre
Nukleinsyrer har en primær, sekundær og tertiær struktur.
Primær struktur av DNA
Den primære strukturen til DNA er en lineær polynukleotidkjede der mononukleotider er forbundet med 3", 5" fosfodiesterbindinger. Utgangsmaterialet for sammenstilling av en nukleinsyrekjede i en celle er 5"-trifosfatnukleosidet, som, som et resultat av fjerning av β og y fosforsyrerester, er i stand til å feste 3" karbonatomet til et annet nukleosid . Således er 3" karbonatomet til en deoksyribose kovalent bundet til 5" karbonatomet til en annen deoksyribose gjennom en enkelt fosforsyrerest og danner en lineær polynukleotidkjede av nukleinsyre. Derav navnet: 3", 5" fosfodiesterbindinger. Nitrogenbaser deltar ikke i å koble sammen nukleotider i én kjede (fig. 1.).
En slik forbindelse, mellom fosforsyremolekylresten til ett nukleotid og karbohydratet til et annet, fører til dannelsen av et pentose-fosfatskjelett av polynukleotidmolekylet, på hvilket nitrogenholdige baser er festet til siden etter hverandre. Deres sekvens av arrangement i kjedene til nukleinsyremolekyler er strengt spesifikke for cellene til forskjellige organismer, dvs. har en bestemt karakter (Chargaffs regel).
En lineær DNA-kjede, hvis lengde avhenger av antall nukleotider inkludert i kjeden, har to ender: den ene kalles 3"-enden og inneholder en fri hydroksyl, og den andre kalles 5"-enden og inneholder en fosforsyre. syrerester. Kretsen er polar og kan ha en retning på 5"->3" og 3"->5". Unntaket er sirkulært DNA.
Den genetiske "teksten" til DNA er sammensatt av kode-"ord" - trillinger av nukleotider kalt kodoner. Deler av DNA som inneholder informasjon om den primære strukturen til alle typer RNA kalles strukturelle gener.
Polynukleotid-DNA-kjeder når gigantiske størrelser, så de pakkes på en bestemt måte i cellen.
Mens han studerte sammensetningen av DNA, etablerte Chargaff (1949) viktige mønstre angående innholdet i individuelle DNA-baser. De hjalp til med å avsløre den sekundære strukturen til DNA. Disse mønstrene kalles Chargaffs regler. Chargaff regler
Disse reglene indikerer at når man konstruerer DNA, må det observeres en ganske streng samsvar (paring) ikke av purin- og pyrimidinbaser generelt, men spesifikt av tymin med adenin og cytosin med guanin. Basert på disse reglene foreslo Watson og Crick i 1953 en modell av den sekundære strukturen til DNA, kalt den doble helixen (fig.). |

Sekundær struktur av DNA
Den sekundære strukturen til DNA er en dobbel helix, modellen som ble foreslått av D. Watson og F. Crick i 1953.
Forutsetninger for å lage en DNA-modell
Som et resultat av innledende analyser ble det antatt at DNA av enhver opprinnelse inneholder alle fire nukleotidene i like molare mengder. På 1940-tallet viste imidlertid E. Chargaff og hans kolleger, som et resultat av å analysere DNA isolert fra en rekke organismer, tydelig at de inneholdt nitrogenholdige baser i forskjellige kvantitative forhold. Chargaff fant at selv om disse forholdene er de samme for DNA fra alle celler av samme organismeart, kan DNA fra forskjellige arter avvike markant i innholdet av visse nukleotider. Dette antydet at forskjellene i forholdet mellom nitrogenholdige baser kan være assosiert med en slags biologisk kode. Selv om forholdet mellom individuelle purin- og pyrimidinbaser i forskjellige DNA-prøver viste seg å være forskjellig, viste det seg et visst mønster ved sammenligning av testresultatene: i alle prøvene var det totale antallet puriner lik det totale antallet pyrimidiner (A + G = T + C), mengden adenin var lik mengden tymin (A = T), og mengden guanin er mengden cytosin (G = C). DNA isolert fra pattedyrceller var generelt rikere på adenin og tymin og relativt fattigere på guanin og cytosin, mens DNA fra bakterier var rikere på guanin og cytosin og relativt fattigere på adenin og tymin. Disse dataene utgjorde en viktig del av faktamaterialet som Watson-Crick-modellen for DNA-struktur senere ble bygget på.
En annen viktig indirekte indikasjon på den mulige strukturen til DNA ble gitt av L. Paulings data om strukturen til proteinmolekyler. Pauling viste at flere forskjellige stabile konfigurasjoner av aminosyrekjeden i et proteinmolekyl er mulig. En vanlig peptidkjedekonfigurasjon, α-helixen, er en vanlig spiralstruktur. Med denne strukturen er dannelsen av hydrogenbindinger mellom aminosyrer lokalisert på tilstøtende svinger av kjeden mulig. Pauling beskrev α-helix-konfigurasjonen av polypeptidkjeden i 1950 og antydet at DNA-molekyler sannsynligvis har en spiralformet struktur holdt på plass av hydrogenbindinger.
Imidlertid ble den mest verdifulle informasjonen om strukturen til DNA-molekylet gitt av resultatene fra røntgendiffraksjonsanalyse. Røntgenstråler som passerer gjennom en DNA-krystall gjennomgår diffraksjon, det vil si at de avbøyes i visse retninger. Graden og arten av avbøyningen av strålene avhenger av strukturen til selve molekylene. Et røntgendiffraksjonsmønster (fig. 3) gir det erfarne øyet en rekke indirekte indikasjoner angående strukturen til molekylene til stoffet som studeres. Analyse av røntgendiffraksjonsmønstre av DNA førte til konklusjonen at de nitrogenholdige basene (som har en flat form) er ordnet som en stabel med plater. Røntgendiffraksjonsmønstre avslørte tre hovedperioder i strukturen til krystallinsk DNA: 0,34, 2 og 3,4 nm.

Watson-Crick DNA-modell
Basert på Chargaffs analytiske data, Wilkins' røntgenmønstre, og forskningen til kjemikere som ga informasjon om de nøyaktige avstandene mellom atomer i et molekyl, vinklene mellom bindingene til et gitt atom og størrelsen på atomene, Watson og Crick begynte å bygge fysiske modeller av de individuelle komponentene i DNA-molekylet i en viss skala og "justere" dem til hverandre på en slik måte at det resulterende systemet tilsvarer forskjellige eksperimentelle data [forestilling] .
Det var kjent enda tidligere at nærliggende nukleotider i en DNA-kjede er forbundet med fosfodiesterbroer, og kobler 5"-karbon deoksyribose-atomet til ett nukleotid med 3"-karbon-deoksyribose-atomet i neste nukleotid. Watson og Crick var ikke i tvil om at perioden på 0,34 nm tilsvarer avstanden mellom suksessive nukleotider i DNA-kjeden. Videre kan det antas at perioden på 2 nm tilsvarer tykkelsen på kjeden. Og for å forklare hvilken reell struktur perioden på 3,4 nm tilsvarer, foreslo Watson og Crick, samt Pauling tidligere, at kjeden er vridd i form av en spiral (eller mer presist, danner en spirallinje, siden en spiral i streng forstand av disse ordene oppnås når spolene danner en konisk snarere enn sylindrisk overflate i rommet). Da vil en periode på 3,4 nm tilsvare avstanden mellom påfølgende svinger av denne helixen. En slik spiral kan være veldig tett eller noe strukket, det vil si at svingene kan være flate eller bratte. Siden perioden på 3,4 nm er nøyaktig 10 ganger avstanden mellom påfølgende nukleotider (0,34 nm), er det klart at hver hel vending av helixen inneholder 10 nukleotider. Fra disse dataene var Watson og Crick i stand til å beregne tettheten til en polynukleotidkjede vridd til en helix med en diameter på 2 nm, med en avstand mellom svingene på 3,4 nm. Det viste seg at en slik kjede ville ha en tetthet som var halvparten av den faktiske tettheten av DNA, som allerede var kjent. Jeg måtte anta at DNA-molekylet består av to kjeder – at det er en dobbel helix av nukleotider.
Den neste oppgaven var selvfølgelig å klargjøre de romlige forholdene mellom de to kjedene som danner den doble helixen. Etter å ha prøvd en rekke alternativer for arrangement av kjeder på deres fysiske modell, fant Watson og Crick at alle tilgjengelige data ble best matchet av alternativet der to polynukleotidhelikser går i motsatte retninger; i dette tilfellet danner kjeder som består av sukker- og fosfatrester overflaten av den doble helixen, og puriner og pyrimidiner er plassert inne. Basene som ligger overfor hverandre, som tilhører to kjeder, er koblet i par med hydrogenbindinger; Det er disse hydrogenbindingene som holder kjedene sammen, og dermed fikserer den generelle konfigurasjonen av molekylet.
Den doble helixen av DNA kan tenkes som en taustige som er vridd på en spiralformet måte, slik at trinnene forblir horisontale. Da vil de to langsgående tauene tilsvare kjeder av sukker- og fosfatrester, og tverrstengene vil tilsvare par av nitrogenholdige baser forbundet med hydrogenbindinger.
Som et resultat av videre studier av mulige modeller, konkluderte Watson og Crick med at hver "tverrstang" skulle bestå av en purin og en pyrimidin; ved en periode på 2 nm (tilsvarende diameteren på dobbelthelixen), ville det ikke være nok plass til to puriner, og de to pyrimidinene kunne ikke være nær nok hverandre til å danne riktige hydrogenbindinger. En dybdestudie av den detaljerte modellen viste at adenin og cytosin, mens de danner en kombinasjon av passende størrelse, fortsatt ikke kunne plasseres på en slik måte at hydrogenbindinger ville dannes mellom dem. Lignende rapporter tvang til å ekskludere kombinasjonen guanin - tymin, mens kombinasjonene adenin - tymin og guanin - cytosin viste seg å være ganske akseptable. Naturen til hydrogenbindinger er slik at adenin danner et par med tymin, og guanin med cytosin. Denne ideen om spesifikk baseparing gjorde det mulig å forklare "Chargaff-regelen", ifølge hvilken mengden adenin i ethvert DNA-molekyl alltid er lik innholdet av tymin, og mengden guanin alltid er lik mengden av cytosin. To hydrogenbindinger dannes mellom adenin og tymin, og tre mellom guanin og cytosin. På grunn av denne spesifisiteten fører dannelsen av hydrogenbindinger mot hvert adenin i en kjede til at tymin dannes på den andre; på samme måte kan bare cytosin være overfor hver guanin. Dermed er kjedene komplementære til hverandre, det vil si at sekvensen av nukleotider i en kjede bestemmer sekvensen deres i den andre. De to kjedene løper i motsatte retninger, og deres terminale fosfatgrupper er i motsatte ender av dobbelthelixen.
Som et resultat av deres forskning foreslo Watson og Crick i 1953 en modell av strukturen til DNA-molekylet (fig. 3), som fortsatt er relevant til i dag. I følge modellen består DNA-molekylet av to komplementære polynukleotidkjeder. Hver DNA-streng er et polynukleotid som består av flere titusenvis av nukleotider. I den danner nabonukleotider en vanlig pentose-fosfat-ryggrad på grunn av koblingen av en fosforsyrerest og deoksyribose med en sterk kovalent binding. De nitrogenholdige basene til en polynukleotidkjede er ordnet i en strengt definert rekkefølge motsatt nitrogenbasene til den andre. Vekslingen av nitrogenholdige baser i en polynukleotidkjede er uregelmessig.
Arrangementet av nitrogenholdige baser i DNA-kjeden er komplementær (fra gresk "komplement" - addisjon), dvs. Tymin (T) er alltid mot adenin (A), og kun cytosin (C) er mot guanin (G). Dette forklares med det faktum at A og T, samt G og C, strengt tatt samsvarer med hverandre, dvs. utfyller hverandre. Denne korrespondansen bestemmes av den kjemiske strukturen til basene, som tillater dannelse av hydrogenbindinger i purin- og pyrimidinparet. Det er to forbindelser mellom A og T, og tre mellom G og C. Disse bindingene gir delvis stabilisering av DNA-molekylet i rommet. Stabiliteten til dobbelthelixen er direkte proporsjonal med antall G≡C-bindinger, som er mer stabile sammenlignet med A=T-bindinger.
Den kjente sekvensen for arrangement av nukleotider i en DNA-kjede gjør det mulig, i henhold til komplementaritetsprinsippet, å etablere nukleotidene til en annen kjede.
I tillegg er det fastslått at nitrogenholdige baser med en aromatisk struktur i en vandig løsning er plassert over hverandre, og danner så å si en stabel med mynter. Denne prosessen med å danne stabler av organiske molekyler kalles stabling. Polynukleotidkjedene til DNA-molekylet til Watson-Crick-modellen som vurderes har en lignende fysisk-kjemisk tilstand, deres nitrogenholdige baser er ordnet i form av en stabel med mynter, mellom planene som van der Waals-interaksjoner (stablingsinteraksjoner) oppstår.
Hydrogenbindinger mellom komplementære baser (horisontalt) og stablingsinteraksjoner mellom plan av baser i en polynukleotidkjede på grunn av van der Waals-krefter (vertikalt) gir DNA-molekylet ytterligere stabilisering i rommet.
Sukkerfosfatryggraden i begge kjedene vender utover, og basene vender innover, mot hverandre. Retningen til kjedene i DNA er antiparallell (en av dem har en retning på 5"->3", den andre - 3"->5", dvs. 3"-enden av en kjede er plassert motsatt 5"-enden av den andre.). Kjedene danner høyrehendte spiraler med felles akse. En omdreining av helixen er 10 nukleotider, størrelsen på svingen er 3,4 nm, høyden på hvert nukleotid er 0,34 nm, diameteren på helixen er 2,0 nm. Som et resultat av rotasjonen av en tråd rundt en annen, dannes et hovedspor (ca. 20 Å i diameter) og et mindre spor (ca. 12 Å i diameter) i DNA-dobbelthelixen. Denne formen av Watson-Crick dobbelthelix ble senere kalt B-formen. I cellene finnes DNA vanligvis i B-form, som er den mest stabile.
Funksjoner av DNA
Den foreslåtte modellen forklarte mange biologiske egenskaper til deoksyribonukleinsyre, inkludert lagring av genetisk informasjon og mangfoldet av gener levert av et bredt utvalg av sekvensielle kombinasjoner av 4 nukleotider og det faktum at det eksisterer en genetisk kode, evnen til å reprodusere seg selv. og overføre genetisk informasjon gitt av replikasjonsprosessen, og implementering av genetisk informasjon i form av proteiner, så vel som andre forbindelser dannet ved hjelp av enzymproteiner.
Grunnleggende funksjoner til DNA.
- DNA er bæreren av genetisk informasjon, som er sikret ved at det finnes en genetisk kode.
- Reproduksjon og overføring av genetisk informasjon på tvers av generasjoner av celler og organismer. Denne funksjonaliteten leveres av replikeringsprosessen.
- Implementering av genetisk informasjon i form av proteiner, samt eventuelle andre forbindelser dannet ved hjelp av enzymproteiner. Denne funksjonen leveres av prosessene med transkripsjon og oversettelse.

Organiseringsformer for dobbelttrådet DNA
DNA kan danne flere typer doble helikser (fig. 4). Foreløpig er seks former allerede kjent (fra A til E og Z-form).
De strukturelle formene for DNA, som Rosalind Franklin etablerte, avhenger av metningen av nukleinsyremolekylet med vann. I studier av DNA-fibre ved bruk av røntgendiffraksjonsanalyse ble det vist at røntgenmønsteret er radikalt avhengig av den relative fuktigheten i hvilken grad av vannmetning av denne fiberen eksperimentet finner sted. Hvis fiberen var tilstrekkelig mettet med vann, ble det tatt ett røntgenbilde. Når den ble tørket, dukket det opp et helt annet røntgenmønster, veldig forskjellig fra røntgenmønsteret til fiber med høy fuktighet.
DNA-molekyl med høy luftfuktighet kalles B-form. Under fysiologiske forhold (lav saltkonsentrasjon, høy grad av hydrering) er den dominerende strukturelle typen DNA B-formen (hovedformen for dobbelttrådet DNA - Watson-Crick-modellen). Helix-stigningen til et slikt molekyl er 3,4 nm. Det er 10 komplementære par per tur i form av vridde stabler av "mynter" - nitrogenholdige baser. Stablene holdes sammen av hydrogenbindinger mellom to motstående "mynter" i stablene, og "vikles" av to bånd av fosfodiester-ryggrad vridd til en høyrehendt helix. Planene til de nitrogenholdige basene er vinkelrett på helixens akse. Tilstøtende komplementære par roteres i forhold til hverandre med 36°. Diameteren på helixen er 20Å, med purin-nukleotidet opptar 12Å og pyrimidin-nukleotidet 8Å.
DNA-molekylet med lavere luftfuktighet kalles A-form. A-formen dannes under forhold med mindre høy hydrering og ved et høyere innhold av Na + eller K + ioner. Denne bredere høyrehendte spiralformede konformasjonen har 11 basepar per tur. Planene til de nitrogenholdige basene har en sterkere helling til helixaksen; de avviker fra normalen til helixaksen med 20°. Dette innebærer tilstedeværelsen av et indre tomrom med en diameter på 5Å. Avstanden mellom tilstøtende nukleotider er 0,23 nm, lengden på svingen er 2,5 nm, og diameteren på helixen er 2,3 nm.
A-formen for DNA ble opprinnelig antatt å være mindre viktig. Imidlertid ble det senere klart at A-formen av DNA, i likhet med B-formen, har enorm biologisk betydning. RNA-DNA-helixen i template-primer-komplekset har A-formen, samt RNA-RNA-helix- og RNA-hårnålsstrukturene (2'-hydroksylgruppen til ribose hindrer RNA-molekyler i å danne B-formen). A-formen av DNA finnes i sporer. Det er fastslått at A-formen av DNA er 10 ganger mer motstandsdyktig mot UV-stråler enn B-formen.
A-formen og B-formen kalles de kanoniske formene for DNA.
Skjemaer C-E også høyrehendt, deres dannelse kan bare observeres i spesielle eksperimenter, og tilsynelatende eksisterer de ikke in vivo. C-formen av DNA har en struktur som ligner på B-DNA. Antall basepar per omdreining er 9,33, lengden på skruevingen er 3,1 nm. Baseparene er skråstilt i en vinkel på 8 grader i forhold til den vinkelrette posisjonen til aksen. Sporene er like i størrelse som sporene til B-DNA. I dette tilfellet er hovedsporet noe grunnere, og det mindre sporet er dypere. Naturlige og syntetiske DNA-polynukleotider kan transformeres til C-formen.
| Tabell 1. Kjennetegn ved noen typer DNA-strukturer | |||
| Spiral type | EN | B | Z |
| Spiral stigning | 0,32 nm | 3,38 nm | 4,46 nm |
| Spiral vri | Ikke sant | Ikke sant | Venstre |
| Antall basepar per tur | 11 | 10 | 12 |
| Avstand mellom grunnplan | 0,256 nm | 0,338 nm | 0,371 nm |
| Konformasjon av glykosidbindinger | anti | anti | anti-C synge |
| Konformasjon av furanoseringen | C3"-endo | C2"-endo | C3"-endo-G C2"-endo-C |
| Sporbredde, liten/stor | 1,11/0,22 nm | 0,57/1,17 nm | 0,2/0,88 nm |
| Rilledybde, liten/stor | 0,26/1,30 nm | 0,82/0,85 nm | 1,38/0,37 nm |
| Spiral diameter | 2,3 nm | 2,0 nm | 1,8 nm |
Strukturelle elementer i DNA
(ikke-kanoniske DNA-strukturer)
De strukturelle elementene i DNA inkluderer uvanlige strukturer begrenset av noen spesielle sekvenser:
|

Z-formet DNA ble oppdaget i 1979 mens han studerte heksanukleotidet d(CG)3 -. Det ble oppdaget av MIT-professor Alexander Rich og hans kolleger. Z-formen har blitt et av de viktigste strukturelle elementene i DNA på grunn av at dannelsen er observert i DNA-regioner der puriner veksler med pyrimidiner (for eksempel 5'-GCGCGC-3'), eller i repetisjoner 5 '-CGCGCG-3' som inneholder metylert cytosin. En vesentlig betingelse for dannelse og stabilisering av Z-DNA var tilstedeværelsen av purin-nukleotider i den i syn-konformasjonen, alternerende med pyrimidinbaser i anti-konformasjonen.
Naturlige DNA-molekyler eksisterer hovedsakelig i høyrehendt B-form med mindre de inneholder sekvenser som (CG)n. Men hvis slike sekvenser er en del av DNA, vil disse seksjonene, når ionestyrken til løsningen eller kationene som nøytraliserer den negative ladningen på fosfodiester-rammeverket endres, kan disse seksjonene transformeres til Z-form, mens andre seksjoner av DNA i kjeden forblir i klassisk B-form. Muligheten for en slik overgang indikerer at de to strengene i DNA-dobbelhelixen er i en dynamisk tilstand og kan slappe av i forhold til hverandre, bevege seg fra den høyrehendte formen til den venstrehendte og vice versa. De biologiske konsekvensene av slik labilitet, som tillater konformasjonstransformasjoner av DNA-strukturen, er ennå ikke fullt ut forstått. Det antas at deler av Z-DNA spiller en viss rolle i å regulere uttrykket av visse gener og deltar i genetisk rekombinasjon.
Z-formen av DNA er en venstrehendt dobbel helix der fosfodiester-ryggraden er plassert i et sikksakkmønster langs molekylets akse. Derav navnet på molekylet (sikksakk)-DNK. Z-DNA er det minst vridde (12 basepar per tur) og tynneste DNA som er kjent i naturen. Avstanden mellom tilstøtende nukleotider er 0,38 nm, lengden på svingen er 4,56 nm, og diameteren til Z-DNA er 1,8 nm. I tillegg er utseendet til dette DNA-molekylet preget av tilstedeværelsen av et enkelt spor.
Z-formen av DNA er funnet i prokaryote og eukaryote celler. Det er nå oppnådd antistoffer som kan skille Z-formen fra B-formen av DNA. Disse antistoffene binder seg til visse områder av de gigantiske kromosomene i spyttkjertelcellene til Drosophila (Dr. melanogaster). Bindingsreaksjonen er lett å overvåke på grunn av den uvanlige strukturen til disse kromosomene, der tettere regioner (disker) står i kontrast til mindre tette regioner (mellomdisker). Z-DNA-regioner er lokalisert i mellomdiskene. Det følger av dette at Z-formen faktisk eksisterer under naturlige forhold, selv om størrelsen på individuelle seksjoner av Z-formen fortsatt er ukjent.

(invertere) er de mest kjente og hyppigst forekommende basesekvensene i DNA. Et palindrom er et ord eller en setning som leser det samme fra venstre til høyre og omvendt. Eksempler på slike ord eller setninger er: HYTE, KOSSAKK, FLOM OG ROSEN FALT PÅ AZORS PÅ. Når det brukes på DNA-seksjoner, betyr dette begrepet (palindrom) den samme vekslingen av nukleotider langs kjeden fra høyre til venstre og venstre til høyre (som bokstavene i ordet "hytte", etc.).
Et palindrom er preget av tilstedeværelsen av inverterte gjentakelser av basesekvenser som har annenordens symmetri i forhold til to DNA-tråder. Slike sekvenser er av åpenbare grunner selvkomplementære og har en tendens til å danne hårnåls- eller korsformede strukturer (fig.). Hårnåler hjelper regulatoriske proteiner med å gjenkjenne hvor den genetiske teksten til kromosom-DNA er kopiert.
Når en omvendt repetisjon er tilstede på den samme DNA-strengen, kalles sekvensen en speilrepetisjon. Speilrepetisjoner har ikke selvkomplementaritetsegenskaper og er derfor ikke i stand til å danne hårnåls- eller korsformede strukturer. Sekvenser av denne typen finnes i nesten alle store DNA-molekyler og kan variere fra bare noen få basepar til flere tusen basepar.
Tilstedeværelsen av palindromer i form av korsformede strukturer i eukaryote celler er ikke påvist, selv om et visst antall korsformede strukturer er påvist in vivo i E. coli-celler. Tilstedeværelsen av selvkomplementære sekvenser i RNA eller enkelttrådet DNA er hovedårsaken til foldingen av nukleinsyrekjeden i løsninger til en viss romlig struktur, preget av dannelsen av mange "hårnåler".

H-form DNA er en helix dannet av tre DNA-tråder - en DNA trippel helix. Det er et kompleks av en Watson-Crick-dobbelthelix med en tredje enkelttrådet DNA-streng, som passer inn i hovedsporet, og danner et såkalt Hoogsteen-par.
Dannelsen av en slik tripleks skjer som et resultat av foldingen av en DNA-dobbelhelix på en slik måte at halvparten av dens seksjon forblir i form av en dobbelhelix, og den andre halvparten er separert. I dette tilfellet danner en av de frakoblede heliksene en ny struktur med den første halvdelen av den doble helixen - en trippel helix, og den andre viser seg å være ustrukturert, i form av en enkeltstrenget seksjon. Et trekk ved denne strukturelle overgangen er dens skarpe avhengighet av pH i mediet, hvis protoner stabiliserer den nye strukturen. På grunn av denne funksjonen ble den nye strukturen kalt H-formen av DNA, hvis dannelse ble oppdaget i supercoiled plasmider som inneholder homopurin-homopyrimidin-regioner, som er en speilrepetisjon.
I ytterligere studier ble det fastslått at det er mulig å utføre en strukturell overgang av noen homopurin-homopyrimidin dobbelttrådet polynukleotider med dannelse av en tre-trådet struktur som inneholder:
- en homopurin og to homopyrimidin tråder ( Py-Pu-Py tripleks) [Hoogsteen-interaksjon].
Konstituerende blokker i Py-Pu-Py-tripleksen er kanoniske isomorfe CGC+- og TAT-triader. Stabilisering av tripleksen krever protonering av CGC+ triaden, så disse tripleksene avhenger av pH i løsningen.
- en homopyrimidin og to homopurintråder ( Py-Pu-Pu tripleks) [omvendt Hoogsteen-interaksjon].
De bestanddelene i Py-Pu-Pu-tripleksen er kanoniske isomorfe CGG- og TAA-triader. En vesentlig egenskap til Py-Pu-Pu-triplekser er avhengigheten av deres stabilitet av tilstedeværelsen av dobbeltladede ioner, og forskjellige ioner kreves for å stabilisere triplekser med forskjellige sekvenser. Siden dannelsen av Py-Pu-Pu-triplekser ikke krever protonering av deres konstituerende nukleotider, kan slike triplekser eksistere ved nøytral pH.
Merk: direkte og omvendte Hoogsteen-interaksjoner forklares av symmetrien til 1-metyltymin: en rotasjon på 180° resulterer i at O2-atomet tar plassen til O4-atomet, mens systemet av hydrogenbindinger er bevart.
To typer trippelhelikser er kjent:
- parallelle trippelhelikser der polariteten til den tredje tråden faller sammen med polariteten til homopurinkjeden til Watson-Crick-dupleksen
- antiparallelle trippelhelikser, der polaritetene til tredje- og homopurinkjedene er motsatte.

G-quadruplex- 4-tråds DNA. Denne strukturen dannes hvis det er fire guaniner, som danner den såkalte G-quadruplex - en runddans på fire guaniner.
De første hintene om muligheten for dannelsen av slike strukturer ble mottatt lenge før gjennombruddsarbeidet til Watson og Crick - tilbake i 1910. Så oppdaget den tyske kjemikeren Ivar Bang at en av komponentene i DNA - guanosinsyre - danner geler i høye konsentrasjoner, mens andre komponenter i DNA ikke har denne egenskapen.
I 1962, ved hjelp av røntgendiffraksjonsmetoden, var det mulig å etablere cellestrukturen til denne gelen. Det viste seg å være sammensatt av fire guaninrester, som forbinder hverandre i en sirkel og danner en karakteristisk firkant. I midten er bindingen støttet av et metallion (Na, K, Mg). De samme strukturene kan dannes i DNA hvis det inneholder mye guanin. Disse flate firkantene (G-kvartettene) er stablet for å danne ganske stabile, tette strukturer (G-quadruplexes).
Fire separate DNA-tråder kan veves inn i fire-trådet komplekser, men dette er snarere et unntak. Oftere er en enkelt tråd av nukleinsyre ganske enkelt knyttet til en knute, og danner karakteristiske fortykkelser (for eksempel ved endene av kromosomer), eller dobbelttrådet DNA i en guaninrik region danner en lokal quadruplex.
Eksistensen av quadruplexes i endene av kromosomer - ved telomerer og i tumorpromotere - har blitt mest studert. Et fullstendig bilde av lokaliseringen av slikt DNA i menneskelige kromosomer er imidlertid fortsatt ikke kjent.
Alle disse uvanlige DNA-strukturene i lineær form er ustabile sammenlignet med B-form DNA. Imidlertid eksisterer DNA ofte i en sirkulær form for topologisk spenning når det har det som kalles supercoiling. Under disse forholdene dannes det lett ikke-kanoniske DNA-strukturer: Z-former, "kryss" og "hårnåler", H-former, guaninkvadruplekser og i-motiv.
- Supercoiled form - notert når den frigjøres fra cellekjernen uten å skade pentosefosfatryggraden. Den har formen av supersnoede lukkede ringer. I supercoil-tilstanden blir DNA-dobbelthelixen "vridd på seg selv" minst én gang, det vil si at den inneholder minst en superturn (tar form av en åttefigur).
- Avslappet tilstand av DNA - observert med et enkelt brudd (brudd på en tråd). I dette tilfellet forsvinner supercoilene og DNA-et tar form av en lukket ring.
- Den lineære formen av DNA observeres når to tråder av en dobbel helix brytes.
Tertiær struktur av DNA
Tertiær struktur av DNA dannes som et resultat av ytterligere vridning i rommet av et dobbeltspiralformet molekyl - dets supercoiling. Supercoiling av DNA-molekylet i eukaryote celler, i motsetning til prokaryoter, skjer i form av komplekser med proteiner.
Nesten alt av DNA fra eukaryoter finnes i kromosomene i kjernene; bare en liten mengde finnes i mitokondrier, og i planter, i plastider. Hovedstoffet i kromosomene til eukaryote celler (inkludert menneskelige kromosomer) er kromatin, bestående av dobbelttrådet DNA, histon og ikke-histonproteiner.
Histonkromatinproteiner
Histoner er enkle proteiner som utgjør opptil 50 % av kromatinet. I alle studerte dyre- og planteceller ble det funnet fem hovedklasser av histoner: H1, H2A, H2B, H3, H4, forskjellig i størrelse, aminosyresammensetning og ladning (alltid positiv).
Pattedyrhiston H1 består av en enkelt polypeptidkjede som inneholder ca. 215 aminosyrer; størrelsen på andre histoner varierer fra 100 til 135 aminosyrer. Alle er spiralisert og vridd til en kule med en diameter på ca. 2,5 nm, og inneholder en uvanlig stor mengde positivt ladede aminosyrer lysin og arginin. Histoner kan være acetylert, metylert, fosforylert, poly(ADP)-ribosylert, og histoner H2A og H2B er kovalent knyttet til ubiquitin. Rollen til slike modifikasjoner i dannelsen av strukturen og ytelsen til funksjoner av histoner har ennå ikke blitt fullstendig belyst. Det antas at dette er deres evne til å samhandle med DNA og gi en av mekanismene for å regulere genvirkning.
Histoner samhandler med DNA primært gjennom ioniske bindinger (saltbroer) dannet mellom de negativt ladede fosfatgruppene i DNA og de positivt ladede lysin- og argininrestene til histoner.
Ikke-histonkromatinproteiner
Ikke-histonproteiner, i motsetning til histoner, er veldig forskjellige. Opptil 590 forskjellige fraksjoner av DNA-bindende ikke-histonproteiner har blitt isolert. De kalles også sure proteiner, siden deres struktur er dominert av sure aminosyrer (de er polyanioner). Mangfoldet av ikke-histonproteiner er assosiert med spesifikk regulering av kromatinaktivitet. For eksempel kan enzymer som kreves for DNA-replikasjon og ekspresjon binde seg til kromatin forbigående. Andre proteiner, f.eks. de som er involvert i forskjellige reguleringsprosesser, binder seg til DNA bare i spesifikt vev eller på visse stadier av differensiering. Hvert protein er komplementært til en spesifikk sekvens av DNA-nukleotider (DNA-sted). Denne gruppen inkluderer:
- familie av stedsspesifikke sinkfingerproteiner. Hver "sinkfinger" gjenkjenner et spesifikt sted som består av 5 nukleotidpar.
- familie av stedspesifikke proteiner - homodimerer. Fragmentet av et slikt protein i kontakt med DNA har en helix-turn-helix-struktur.
- høymobilitetsgelproteiner (HMG-proteiner) er en gruppe strukturelle og regulatoriske proteiner som er konstant assosiert med kromatin. De har en molekylvekt på mindre enn 30 kDa og er preget av et høyt innhold av ladede aminosyrer. På grunn av deres lave molekylvekt har HMG-proteiner høy mobilitet under polyakrylamidgelelektroforese.
- replikering, transkripsjon og reparasjonsenzymer.
Med deltakelse av strukturelle, regulatoriske proteiner og enzymer involvert i syntesen av DNA og RNA, omdannes nukleosomtråden til et svært kondensert kompleks av proteiner og DNA. Den resulterende strukturen er 10 000 ganger kortere enn det opprinnelige DNA-molekylet.

Kromatin
Kromatin er et kompleks av proteiner med kjernefysisk DNA og uorganiske stoffer. Hoveddelen av kromatinet er inaktivt. Den inneholder tettpakket, kondensert DNA. Dette er heterochromatin. Det er konstitutivt, genetisk inaktivt kromatin (satellitt-DNA) som består av ikke-uttrykte områder, og fakultativt - inaktivt i en rekke generasjoner, men under visse omstendigheter i stand til å uttrykkes.
Aktivt kromatin (eukromatin) er ukondensert, dvs. pakket mindre tett. I forskjellige celler varierer innholdet fra 2 til 11%. I hjerneceller er det mest rikelig - 10-11%, i leverceller - 3-4 og nyreceller - 2-3%. Aktiv transkripsjon av eukromatin er notert. Dessuten tillater dens strukturelle organisering at den samme genetiske DNA-informasjonen som er iboende i en gitt type organisme, kan brukes annerledes i spesialiserte celler.
I et elektronmikroskop ligner bildet av kromatin på perler: sfæriske fortykkelser omtrent 10 nm store, adskilt av trådlignende broer. Disse sfæriske fortykningene kalles nukleosomer. Nukleosomet er en strukturell enhet av kromatin. Hvert nukleosom inneholder et 146-bp supercoiled DNA-segment sår for å danne 1,75 venstre svinger per nukleosomal kjerne. Den nukleosomale kjernen er en histonoktamer bestående av histonene H2A, H2B, H3 og H4, to molekyler av hver type (fig. 9), som ser ut som en skive med en diameter på 11 nm og en tykkelse på 5,7 nm. Den femte histonen, H1, er ikke en del av den nukleosomale kjernen og er ikke involvert i prosessen med å vikle DNA til histonoktameren. Den kontakter DNA på stedene der den doble helixen kommer inn og ut av den nukleosomale kjernen. Dette er intercore (linker) DNA-seksjoner, hvor lengden varierer avhengig av celletypen fra 40 til 50 nukleotidpar. Som et resultat varierer også lengden på DNA-fragmentet inkludert i nukleosomene (fra 186 til 196 nukleotidpar).
Nukleosomer inneholder omtrent 90% DNA, resten er linkere. Det antas at nukleosomer er fragmenter av "stille" kromatin, og linkeren er aktiv. Imidlertid kan nukleosomer utfolde seg og bli lineære. Ufoldede nukleosomer er allerede aktive kromatin. Dette viser tydelig funksjonens avhengighet av struktur. Det kan antas at jo mer kromatin er inneholdt i kuleformede nukleosomer, jo mindre aktivt er det. Det er klart at i forskjellige celler er den ulik andel hvilekromatin assosiert med antallet slike nukleosomer.
I elektronmikroskopiske fotografier, avhengig av isolasjonsforholdene og graden av strekking, kan kromatin ikke bare se ut som en lang tråd med fortykkelser - "perler" av nukleosomer, men også som en kortere og tettere fibril (fibril) med en diameter på 30 nm, hvis dannelse observeres under interaksjon histon H1 bundet til linkerregionen til DNA og histon H3, noe som fører til ytterligere vridning av helixen på seks nukleosomer per omdreining for å danne en solenoid med en diameter på 30 nm. I dette tilfellet kan histonproteinet forstyrre transkripsjonen av en rekke gener og dermed regulere deres aktivitet.
Som et resultat av interaksjonene mellom DNA og histoner beskrevet ovenfor, omdannes et segment av en DNA-dobbelthelix på 186 basepar med en gjennomsnittlig diameter på 2 nm og en lengde på 57 nm til en helix med en diameter på 10 nm og en lengde på 5 nm. Når denne spiralen deretter komprimeres til en fiber med en diameter på 30 nm, øker graden av kondensering ytterligere seks ganger.
Til syvende og sist resulterer pakkingen av en DNA-dupleks med fem histoner i 50 ganger kondensering av DNA. Selv en så høy grad av kondensering kan imidlertid ikke forklare den nesten 50 000 - 100 000 ganger komprimeringen av DNA i metafasekromosomet. Dessverre er detaljene for ytterligere kromatinpakking opp til metafasekromosomet ennå ikke kjent, så vi kan bare vurdere de generelle egenskapene til denne prosessen.
Nivåer av DNA-komprimering i kromosomer
Hvert DNA-molekyl er pakket inn i et separat kromosom. Humane diploide celler inneholder 46 kromosomer, som er lokalisert i cellekjernen. Den totale lengden på DNAet til alle kromosomene i en celle er 1,74 m, men diameteren på kjernen som kromosomene er pakket i er millioner av ganger mindre. Slik kompakt pakking av DNA i kromosomer og kromosomer i cellekjernen sikres av en rekke histon- og ikke-histonproteiner som interagerer i en bestemt sekvens med DNA (se ovenfor). Komprimering av DNA i kromosomer gjør det mulig å redusere dens lineære dimensjoner med omtrent 10 000 ganger - omtrent fra 5 cm til 5 mikron. Det er flere nivåer av komprimering (fig. 10).


- DNA dobbel helix er et negativt ladet molekyl med en diameter på 2 nm og en lengde på flere cm.
- nukleosomnivå- kromatin ser i et elektronmikroskop ut som en kjede av "perler" - nukleosomer - "på en tråd". Nukleosomet er en universell strukturell enhet som finnes i både eukromatin og heterokromatin, i interfasekjernen og metafasekromosomene.
Det nukleosomale nivået av komprimering er sikret av spesielle proteiner - histoner. Åtte positivt ladede histondomener danner kjernen i nukleosomet som et negativt ladet DNA-molekyl er viklet rundt. Dette gir en avkortning på 7 ganger, mens diameteren øker fra 2 til 11 nm.
- solenoidnivå
Solenoidnivået til kromosomorganisering er preget av vridning av nukleosomfilamentet og dannelsen av tykkere fibriller med en diameter på 20-35 nm - solenoider eller superbider. Solenoidstigningen er 11 nm; det er omtrent 6-10 nukleosomer per omdreining. Solenoidpakking anses som mer sannsynlig enn superbid pakking, ifølge hvilken en kromatinfibril med en diameter på 20-35 nm er en kjede av granuler, eller superbids, som hver består av åtte nukleosomer. På solenoidnivå reduseres den lineære størrelsen på DNA med 6-10 ganger, diameteren øker til 30 nm.
- løkkenivå
Løkkenivået er gitt av ikke-histon-stedspesifikke DNA-bindende proteiner som gjenkjenner og binder seg til spesifikke DNA-sekvenser, og danner løkker på omtrent 30-300 kb. Sløyfen sikrer genuttrykk, d.v.s. løkken er ikke bare en strukturell, men også en funksjonell formasjon. Forkorting på dette nivået skjer 20-30 ganger. Diameteren øker til 300 nm. Sløyfeformede strukturer som «lampebørster» i amfibieoocytter kan sees i cytologiske preparater. Disse løkkene ser ut til å være supercoiled og representerer DNA-domener, sannsynligvis tilsvarende enheter for transkripsjon og kromatinreplikasjon. Spesifikke proteiner fikserer basene til løkkene og muligens noen av deres indre seksjoner. Den løkkelignende domeneorganisasjonen fremmer folding av kromatin i metafasekromosomer til spiralformede strukturer av høyere orden.
- domenenivå
Domenenivået for kromosomorganisering er ikke studert nok. På dette nivået noteres dannelsen av løkkedomener - strukturer av tråder (fibriller) 25-30 nm tykke, som inneholder 60% protein, 35% DNA og 5% RNA, er praktisk talt usynlige i alle faser av cellesyklusen med unntak av mitose og er noe tilfeldig fordelt gjennom cellekjernen. Sløyfeformede strukturer som «lampebørster» i amfibieoocytter kan sees i cytologiske preparater.
Loop-domener er festet ved basen til den intranukleære proteinmatrisen i de såkalte innebygde festestedene, ofte referert til som MAR/SAR-sekvenser (MAR, fra den engelske matriseassosierte regionen; SAR, fra de engelske stillasfesteregionene) - DNA-fragmenter flere hundre lange basepar som er karakterisert ved et høyt innhold (>65%) av A/T-nukleotidpar. Hvert domene ser ut til å ha et enkelt replikasjonsopphav og fungerer som en autonom superhelisk enhet. Ethvert løkkedomene inneholder mange transkripsjonsenheter, hvis funksjon sannsynligvis er koordinert - hele domenet er enten i en aktiv eller inaktiv tilstand.
På domenenivå, som et resultat av sekvensiell kromatinpakking, skjer en reduksjon i de lineære dimensjonene til DNA med omtrent 200 ganger (700 nm).
- kromosomnivå
På kromosomnivå skjer kondensering av profasekromosomet til et metafasekromosom med komprimering av løkkedomener rundt det aksiale rammeverket til ikke-histonproteiner. Denne supercoilingen er ledsaget av fosforylering av alle H1-molekyler i cellen. Som et resultat kan metafasekromosomet avbildes som tettpakkede solenoidsløyfer, kveilet inn i en tett spiral. Et typisk menneskelig kromosom kan inneholde opptil 2600 løkker. Tykkelsen på en slik struktur når 1400 nm (to kromatider), og DNA-molekylet er forkortet med 104 ganger, dvs. fra 5 cm strukket DNA til 5 µm.
Funksjoner av kromosomer
I samspill med ekstrakromosomale mekanismer gir kromosomer
- lagring av arvelig informasjon
- bruke denne informasjonen til å opprette og vedlikeholde mobilorganisasjon
- regulering av lesing av arvelig informasjon
- selvduplisering av genetisk materiale
- overføring av genetisk materiale fra modercellen til dattercellene.
Det er bevis på at når en region av kromatin aktiveres, dvs. under transkripsjon fjernes først histon H1 og deretter histonoktetten reversibelt fra den. Dette forårsaker kromatin-dekondensering, den sekvensielle overgangen av en 30-nm kromatinfibril til en 10-nm-fibril og dens videre utfolding til seksjoner av fritt DNA, dvs. tap av nukleosomstruktur.
Etter oppdagelsen av prinsippet om molekylær organisering av et slikt stoff som DNA i 1953, begynte molekylærbiologi å utvikle seg. Videre i forskningsprosessen fant forskerne ut hvordan DNA er rekombinert, dets sammensetning og hvordan vårt menneskelige genom er strukturert.
Hver dag skjer komplekse prosesser på molekylært nivå. Hvordan er DNA-molekylet bygget opp, hva består det av? Og hvilken rolle spiller DNA-molekyler i en celle? La oss snakke i detalj om alle prosessene som skjer inne i dobbeltkjeden.
Hva er arvelig informasjon?
Så hvor begynte det hele? Tilbake i 1868 fant de det i bakteriekjernene. Og i 1928 fremmet N. Koltsov teorien om at all genetisk informasjon om en levende organisme er kryptert i DNA. Så fant J. Watson og F. Crick en modell for den nå velkjente DNA-helixen i 1953, som de fortjent fikk anerkjennelse og en pris for – Nobelprisen.
Hva er DNA egentlig? Dette stoffet består av 2 forente tråder, eller snarere spiraler. En del av en slik kjede med viss informasjon kalles et gen.

DNA lagrer all informasjon om hva slags proteiner som vil bli dannet og i hvilken rekkefølge. DNA-makromolekylet er en materiell bærer av utrolig voluminøs informasjon, som er registrert i en streng sekvens av individuelle murstein - nukleotider. Det er totalt 4 nukleotider; de utfyller hverandre kjemisk og geometrisk. Dette prinsippet om komplementaritet, eller komplementaritet, i vitenskapen vil bli beskrevet senere. Denne regelen spiller en nøkkelrolle i koding og dekoding av genetisk informasjon.
Siden DNA-tråden er utrolig lang, er det ingen repetisjoner i denne sekvensen. Hver levende skapning har sin egen unike DNA-streng.
Funksjoner av DNA
Funksjoner inkluderer lagring av arvelig informasjon og overføring av den til avkom. Uten denne funksjonen kunne genomet til en art ikke bevares og utvikles over tusenvis av år. Organismer som har gjennomgått alvorlige genmutasjoner er mer sannsynlig å ikke overleve eller miste evnen til å produsere avkom. Slik oppstår naturlig beskyttelse mot degenerasjonen av arten.

En annen viktig funksjon er implementering av lagret informasjon. En celle kan ikke lage et enkelt viktig protein uten de instruksjonene som er lagret i en dobbelkjede.
Nukleinsyresammensetning
Det er nå kjent med sikkerhet hva selve nukleotidene – byggesteinene til DNA – er laget av. De inneholder 3 stoffer:
- Ortofosforsyre.
- Nitrogenholdig base. Pyrimidinbaser - som kun har en ring. Disse inkluderer tymin og cytosin. Purinbaser, som inneholder 2 ringer. Disse er guanin og adenin.
- sukrose. DNA inneholder deoksyribose, RNA inneholder ribose.
Antall nukleotider er alltid lik antall nitrogenholdige baser. I spesielle laboratorier brytes nukleotidet ned og nitrogenbasen isoleres fra det. Slik studeres de individuelle egenskapene til disse nukleotidene og mulige mutasjoner i dem.
Nivåer for organisering av arvelig informasjon
Det er 3 nivåer av organisering: genetisk, kromosomal og genomisk. All informasjon som trengs for syntesen av et nytt protein finnes i en liten del av kjeden - genet. Det vil si at genet anses som det laveste og enkleste nivået av informasjonskoding.

Gener på sin side er satt sammen til kromosomer. Takket være denne organisasjonen av bæreren av arvelig materiale, veksler grupper av egenskaper i henhold til visse lover og overføres fra en generasjon til en annen. Det skal bemerkes at det er utrolig mange gener i kroppen, men informasjonen går ikke tapt selv når den rekombineres mange ganger.
Det finnes flere typer gener:
- I henhold til deres funksjonelle formål er det 2 typer: strukturelle og regulatoriske sekvenser;
- Basert på deres innflytelse på prosessene som skjer i cellen, skiller de: supervitale, dødelige, betinget dødelige gener, samt mutator- og antimutatorgener.
Gener er ordnet langs kromosomet i en lineær rekkefølge. I kromosomer er informasjon ikke fokusert tilfeldig; det er en viss rekkefølge. Det er til og med et kart som viser posisjonene, eller lokiene, til gener. For eksempel er det kjent at kromosom nr. 18 krypterer data om fargen på et barns øyne.
Hva er et genom? Dette er navnet gitt til hele settet med nukleotidsekvenser i en organismes celle. Genomet karakteriserer en hel art, ikke et individ.
Hva er menneskets genetiske kode?
Faktum er at hele det enorme potensialet for menneskelig utvikling ligger allerede i unnfangelsesperioden. All arvelig informasjon som er nødvendig for utviklingen av zygoten og veksten av barnet etter fødselen er kryptert i gener. DNA-seksjoner er de mest grunnleggende bærerne av arvelig informasjon.
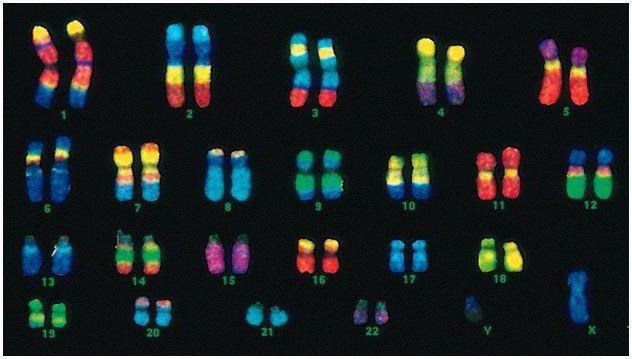
Mennesker har 46 kromosomer, eller 22 somatiske par pluss ett kjønnsbestemmende kromosom fra hver forelder. Dette diploide settet med kromosomer koder for hele det fysiske utseendet til en person, hans mentale og fysiske evner og mottakelighet for sykdommer. Somatiske kromosomer er utad ikke å skille, men de har forskjellig informasjon, siden en av dem er fra faren, den andre fra moren.
Den mannlige koden skiller seg fra den kvinnelige koden i det siste kromosomparet - XY. Det kvinnelige diploide settet er det siste paret, XX. Hannene får ett X-kromosom fra sin biologiske mor, som deretter blir gitt videre til døtrene deres. Kjønns Y-kromosomet overføres til sønner.
Menneskelige kromosomer varierer sterkt i størrelse. For eksempel er det minste kromosomparet nr. 17. Og det største paret er 1 og 3.
Diameteren på dobbeltspiralen hos mennesker er bare 2 nm. DNA-et er kveilet så tett at det passer inn i den lille cellekjernen, selv om det ville vært opptil 2 meter langt hvis det ikke ble vridd. Lengden på helixen er hundrevis av millioner nukleotider.
Hvordan overføres den genetiske koden?
Så hvilken rolle spiller DNA-molekyler i celledeling? Gener - bærere av arvelig informasjon - er plassert inne i hver celle i kroppen. For å videreformidle koden sin til en datterorganisme deler mange skapninger sitt DNA i 2 identiske helikser. Dette kalles replikering. Under replikasjonsprosessen vikles DNA av og spesielle "maskiner" fullfører hver tråd. Etter at den genetiske helixen deler seg, begynner kjernen og alle organeller å dele seg, og deretter hele cellen.
Men mennesker har en annen prosess for genoverføring - seksuell. Karakteristikkene til far og mor er blandet, den nye genetiske koden inneholder informasjon fra begge foreldrene.
Lagring og overføring av arvelig informasjon er mulig på grunn av den komplekse organiseringen av DNA-helixen. Tross alt, som vi sa, er strukturen til proteiner kryptert i gener. Når den er opprettet på unnfangelsestidspunktet, vil denne koden kopiere seg selv gjennom hele livet. Karyotypen (personlig sett med kromosomer) endres ikke under fornyelsen av organceller. Overføringen av informasjon utføres ved hjelp av kjønnskjønnsceller - mannlige og kvinnelige.
Bare virus som inneholder én RNA-streng er ikke i stand til å overføre informasjonen deres til deres avkom. Derfor trenger de menneske- eller dyreceller for å reprodusere seg.
Implementering av arvelig informasjon
Viktige prosesser skjer hele tiden i cellekjernen. All informasjon registrert i kromosomer brukes til å bygge proteiner fra aminosyrer. Men DNA-kjeden forlater aldri kjernen, så den trenger hjelp av en annen viktig forbindelse: RNA. Det er RNA som er i stand til å trenge gjennom kjernemembranen og samhandle med DNA-kjeden.
Gjennom samspillet mellom DNA og 3 typer RNA blir all kodet informasjon realisert. På hvilket nivå skjer implementering av arvelig informasjon? Alle interaksjoner skjer på nukleotidnivå. Messenger RNA kopierer en del av DNA-kjeden og bringer denne kopien til ribosomet. Her begynner syntesen av et nytt molekyl fra nukleotider.
For at mRNA skal kopiere den nødvendige delen av kjeden, utfolder helixen seg og gjenopprettes deretter, etter fullføring av omkodingsprosessen. Dessuten kan denne prosessen skje samtidig på 2 sider av 1 kromosom.
Prinsippet om komplementaritet
De består av 4 nukleotider - adenin (A), guanin (G), cytosin (C), tymin (T). De er forbundet med hydrogenbindinger i henhold til regelen om komplementaritet. Arbeidet til E. Chargaff bidro til å etablere denne regelen, siden forskeren la merke til noen mønstre i oppførselen til disse stoffene. E. Chargaff oppdaget at molforholdet mellom adenin og tymin er lik én. Og på samme måte er forholdet mellom guanin og cytosin alltid lik én.
Basert på hans arbeid dannet genetikere en regel for samspillet mellom nukleotider. Komplementaritetsregelen sier at adenin bare kombineres med tymin, og guanin bare kombineres med cytosin. Under avkodingen av helixen og syntesen av et nytt protein i ribosomet, hjelper denne vekslingsregelen til raskt å finne den nødvendige aminosyren som er festet til overførings-RNA.
RNA og dets typer
Hva er arvelig informasjon? nukleotider i en dobbel DNA-streng. Hva er RNA? Hva er jobben hennes? RNA, eller ribonukleinsyre, hjelper til med å trekke ut informasjon fra DNA, dekode den og, basert på komplementaritetsprinsippet, skape proteiner som er nødvendige for celler.
Det er 3 typer RNA totalt. Hver av dem utfører strengt tatt sin egen funksjon.
- Informasjonsinformasjon (mRNA), eller også kalt matrise. Den går rett inn i midten av cellen, inn i kjernen. Finner i et av kromosomene det nødvendige genetiske materialet for å bygge et protein og kopierer en av sidene av dobbelttråden. Kopiering skjer igjen i henhold til komplementaritetsprinsippet.
- Transportere er et lite molekyl som har nukleotiddekodere på den ene siden, og aminosyrer som tilsvarer grunnkoden på den andre siden. Oppgaven til tRNA er å levere det til "verkstedet", det vil si til ribosomet, hvor det syntetiserer den nødvendige aminosyren.
- rRNA er ribosomalt. Den kontrollerer mengden protein som produseres. Den består av 2 deler - en aminosyre og en peptiddel.
Den eneste forskjellen i dekoding er at RNA ikke har tymin. I stedet for tymin er uracil tilstede her. Men så, under prosessen med proteinsyntese, installerer tRNA fortsatt alle aminosyrene riktig. Hvis det oppstår feil i dekodingsinformasjonen, oppstår en mutasjon.
Reparasjon av skadet DNA-molekyl
Prosessen med å gjenopprette en skadet dobbeltstreng kalles reparasjon. Under reparasjonsprosessen fjernes skadede gener.

Deretter reproduseres den nødvendige sekvensen av elementer nøyaktig og kuttes tilbake til samme sted på kjeden der den ble fjernet. Alt dette skjer takket være spesielle kjemikalier - enzymer.
Hvorfor oppstår mutasjoner?
Hvorfor begynner noen gener å mutere og slutter å utføre sin funksjon - lagre viktig arvelig informasjon? Dette oppstår på grunn av en feil i dekodingen. For eksempel hvis adenin ved et uhell erstattes med tymin.
Det er også kromosomale og genomiske mutasjoner. Kromosomale mutasjoner oppstår når deler av arvelig informasjon går tapt, dupliseres eller til og med overføres og settes inn i et annet kromosom.

Genomiske mutasjoner er de mest alvorlige. Årsaken deres er en endring i antall kromosomer. Det vil si når i stedet for et par - et diploid sett, er et triploid sett til stede i karyotypen.
Det mest kjente eksemplet på en triploid mutasjon er Downs syndrom, der det personlige settet med kromosomer er 47. Hos slike barn dannes 3 kromosomer i stedet for det 21. paret.
Det er også en kjent mutasjon som kalles polyploidi. Men polyploidi forekommer bare i planter.
DNA-molekylet består av to tråder som danner en dobbel helix. Strukturen ble først dechiffrert av Francis Crick og James Watson i 1953.
Først ga DNA-molekylet, bestående av et par nukleotidkjeder vridd rundt hverandre, opphav til spørsmål om hvorfor det hadde akkurat denne formen. Forskere kaller dette fenomenet komplementaritet, som betyr at bare visse nukleotider kan bli funnet overfor hverandre i trådene. For eksempel er adenin alltid motsatt tymin, og guanin er alltid motsatt cytosin. Disse nukleotidene til DNA-molekylet kalles komplementære.
Skjematisk er det avbildet slik:
T - A
C - G
Disse parene danner en kjemisk nukleotidbinding, som bestemmer rekkefølgen av aminosyrer. I det første tilfellet er det litt svakere. Forbindelsen mellom C og G er sterkere. Ikke-komplementære nukleotider danner ikke par med hverandre.
Om bygningen
Så strukturen til DNA-molekylet er spesiell. Den har denne formen av en grunn: faktum er at antallet nukleotider er veldig stort, og det trengs mye plass for å romme lange kjeder. Det er av denne grunn at kjedene er preget av en spiralvridning. Dette fenomenet kalles spiralisering, det lar trådene forkortes med omtrent fem til seks ganger.
Kroppen bruker noen molekyler av denne typen veldig aktivt, andre sjelden. Sistnevnte, i tillegg til spiralisering, gjennomgår også en slik "kompakt emballasje" som superspiralisering. Og så minker lengden på DNA-molekylet med 25-30 ganger.

Hva er "emballasjen" til et molekyl?
Prosessen med supercoiling involverer histonproteiner. De har strukturen og utseendet til en trådsnelle eller en stang. Spiraliserte tråder vikles på dem, som umiddelbart blir "kompakt pakket" og tar liten plass. Når det oppstår behov for å bruke en eller annen tråd, vikles den av en spole, for eksempel et histonprotein, og spiralen vikles av i to parallelle kjeder. Når DNA-molekylet er i denne tilstanden, kan de nødvendige genetiske dataene leses fra det. Det er imidlertid én betingelse. Innhenting av informasjon er bare mulig hvis strukturen til DNA-molekylet har en ikke-vridd form. Kromosomer som er tilgjengelige for lesing kalles eukromatiner, og hvis de er supercoiled, er de allerede heterokromatiner.
Nukleinsyrer
Nukleinsyrer, som proteiner, er biopolymerer. Hovedfunksjonen er lagring, implementering og overføring av arvelig (genetisk informasjon). De kommer i to typer: DNA og RNA (deoksyribonuklein og ribonuklein). Monomerene i dem er nukleotider, som hver inneholder en fosforsyrerest, et femkarbonsukker (deoksyribose/ribose) og en nitrogenholdig base. DNA-koden inkluderer 4 typer nukleotider - adenin (A) / guanin (G) / cytosin (C) / tymin (T). De er forskjellige i den nitrogenholdige basen de inneholder.
I et DNA-molekyl kan antallet nukleotider være enormt – fra flere tusen til titalls og hundrevis av millioner. Slike gigantiske molekyler kan undersøkes gjennom et elektronmikroskop. I dette tilfellet vil du kunne se en dobbel kjede av polynukleotidtråder, som er forbundet med hverandre ved hjelp av hydrogenbindinger av de nitrogenholdige basene til nukleotidene.

Forskning
I løpet av forskningen oppdaget forskere at typene DNA-molekyler er forskjellige i forskjellige levende organismer. Det ble også funnet at guanin i en kjede bare kan binde seg til cytosin, og tymin til adenin. Arrangementet av nukleotider i en kjede tilsvarer strengt tatt den parallelle. Takket være denne komplementariteten til polynukleotider, er DNA-molekylet i stand til å dobles og selvreproduksjon. Men først divergerer de komplementære kjedene, under påvirkning av spesielle enzymer som ødelegger sammenkoblede nukleotider, og deretter begynner syntesen av den manglende kjeden i hver av dem. Dette skjer på grunn av de frie nukleotidene som er tilstede i store mengder i hver celle. Som et resultat av dette, i stedet for "modermolekylet", dannes to "datter" som er identiske i sammensetning og struktur, og DNA-koden blir den opprinnelige. Denne prosessen er en forløper til celledeling. Det sikrer overføring av alle arvelige data fra morceller til datterceller, så vel som til alle påfølgende generasjoner.
Hvordan leses genkoden?
I dag beregnes ikke bare massen til et DNA-molekyl – det er også mulig å finne ut mer komplekse data som tidligere var utilgjengelige for forskere. Du kan for eksempel lese informasjon om hvordan en organisme bruker sin egen celle. Selvfølgelig er denne informasjonen først i kodet form og har form av en viss matrise, og derfor må den transporteres til en spesiell bærer, som er RNA. Ribonukleinsyre er i stand til å trenge inn i cellen gjennom kjernemembranen og lese den kodede informasjonen inni. Dermed er RNA en bærer av skjulte data fra kjernen til cellen, og det skiller seg fra DNA ved at det inneholder ribose i stedet for deoksyribose, og uracil i stedet for tymin. I tillegg er RNA enkelttrådet.

RNA syntese
Dybdeanalyse av DNA har vist at etter at RNA forlater kjernen, kommer det inn i cytoplasmaet, hvor det kan integreres som en matrise i ribosomer (spesielle enzymsystemer). Veiledet av informasjonen som mottas, kan de syntetisere den passende sekvensen av proteinaminosyrer. Ribosomet lærer av triplettkoden hvilken type organisk forbindelse som må festes til den dannede proteinkjeden. Hver aminosyre har sin egen spesifikke triplett, som koder for den.
Etter at dannelsen av kjeden er fullført, får den en spesifikk romlig form og blir til et protein som er i stand til å utføre sine hormonelle, konstruksjonsmessige, enzymatiske og andre funksjoner. For enhver organisme er det et genprodukt. Det er fra det at alle slags egenskaper, egenskaper og manifestasjoner av gener bestemmes.

Gener
Sekvenseringsprosesser ble først og fremst utviklet for å få informasjon om hvor mange gener et DNA-molekyl har i sin struktur. Og selv om forskning har gjort det mulig for forskere å gjøre store fremskritt i denne saken, er det ennå ikke mulig å vite det nøyaktige antallet.
For bare noen få år siden ble det antatt at DNA-molekyler inneholder omtrent 100 tusen gener. Litt senere sank tallet til 80 tusen, og i 1998 uttalte genetikere at bare 50 tusen gener er til stede i ett DNA, som bare er 3% av den totale DNA-lengden. Men de siste konklusjonene til genetikere var slående. Nå hevder de at genomet inkluderer 25-40 tusen av disse enhetene. Det viser seg at bare 1,5 % av kromosomalt DNA er ansvarlig for koding av proteiner.
Forskningen stoppet ikke der. Et parallelt team av genteknologispesialister fant at antallet gener i ett molekyl er nøyaktig 32 tusen. Som du kan se, er det fortsatt umulig å få et definitivt svar. Det er for mange motsetninger. Alle forskere stoler kun på resultatene sine.
Var det evolusjon?
Til tross for at det ikke er noen bevis for utviklingen av molekylet (siden strukturen til DNA-molekylet er skjør og liten i størrelse), gjorde forskerne fortsatt en antagelse. Basert på laboratoriedata ga de uttrykk for følgende versjon: i den innledende fasen av utseendet hadde molekylet form av et enkelt selvreplikerende peptid, som inkluderte opptil 32 aminosyrer funnet i de gamle havene.
Etter selvreplikasjon, takket være kreftene til naturlig utvalg, fikk molekyler evnen til å beskytte seg mot ytre elementer. De begynte å leve lenger og reprodusere seg i større mengder. Molekyler som befant seg i lipidboblen hadde alle muligheter til å reprodusere seg selv. Som et resultat av en rekke påfølgende sykluser, fikk lipidbobler form av cellemembraner, og deretter - de velkjente partiklene. Det skal bemerkes at i dag er enhver del av et DNA-molekyl en kompleks og tydelig fungerende struktur, alle funksjonene som forskerne ennå ikke har studert fullt ut.

Moderne verden
Nylig har forskere fra Israel utviklet en datamaskin som kan utføre billioner av operasjoner per sekund. I dag er det den raskeste bilen på jorden. Hele hemmeligheten er at den innovative enheten er drevet av DNA. Professorer sier at i nær fremtid vil slike datamaskiner til og med kunne generere energi.
For et år siden kunngjorde spesialister fra Weizmann Institute i Rehovot (Israel) etableringen av en programmerbar molekylær datamaskin bestående av molekyler og enzymer. De byttet ut silisiummikrobrikker med dem. Til dags dato har teamet gjort ytterligere fremgang. Nå kan bare ett DNA-molekyl gi en datamaskin de nødvendige dataene og det nødvendige drivstoffet.
Biokjemiske "nanodatamaskiner" er ikke en fiksjon; de eksisterer allerede i naturen og er manifestert i enhver levende skapning. Men ofte blir de ikke administrert av mennesker. En person kan ennå ikke operere på genomet til noen plante for å beregne for eksempel tallet "Pi".
Ideen om å bruke DNA til å lagre/behandle data kom først i hodet til forskere i 1994. Det var da et molekyl ble brukt til å løse et enkelt matematisk problem. Siden den gang har en rekke forskergrupper foreslått ulike prosjekter knyttet til DNA-datamaskiner. Men her var alle forsøk kun basert på energimolekylet. Du kan ikke se en slik datamaskin med det blotte øye, den ser ut som en gjennomsiktig løsning av vann i et reagensrør. Det er ingen mekaniske deler i den, men bare billioner av biomolekylære enheter - og dette er bare i en dråpe væske!
Menneskelig DNA
Folk ble klar over typen menneskelig DNA i 1953, da forskerne først var i stand til å demonstrere for verden en dobbelttrådet DNA-modell. For dette fikk Kirk og Watson Nobelprisen, siden denne oppdagelsen ble grunnleggende på 1900-tallet.
Over tid beviste de selvfølgelig at et strukturert menneskelig molekyl ikke bare kan se ut som i den foreslåtte versjonen. Etter å ha utført en mer detaljert DNA-analyse, oppdaget de A-, B- og venstrehendt form Z-. Form A- er ofte et unntak, siden den dannes bare hvis det er mangel på fuktighet. Men dette er bare mulig i laboratoriestudier; for det naturlige miljøet er dette unormalt; en slik prosess kan ikke forekomme i en levende celle.
B-formen er klassisk og er kjent som en dobbel høyrehendt kjede, men Z-formen er ikke bare vridd i motsatt retning til venstre, men har også et mer sikksakk-utseende. Forskere har også identifisert G-quadruplex-formen. Strukturen har ikke 2, men 4 tråder. Ifølge genetikere forekommer denne formen i områder hvor det er en overflødig mengde guanin.

Kunstig DNA
I dag finnes det allerede kunstig DNA, som er en identisk kopi av det ekte; den følger perfekt strukturen til den naturlige doble helixen. Men i motsetning til det opprinnelige polynukleotidet, har det kunstige bare to ekstra nukleotider.
Siden dubbingen ble laget basert på informasjon hentet fra ulike studier av ekte DNA, kan den også kopieres, selvreplikeres og utvikles. Eksperter har jobbet med å lage et slikt kunstig molekyl i omtrent 20 år. Resultatet er en fantastisk oppfinnelse som kan bruke den genetiske koden på samme måte som naturlig DNA.
Til de fire eksisterende nitrogenholdige basene la genetikere ytterligere to, som ble opprettet ved kjemisk modifisering av naturlige baser. I motsetning til naturlig DNA, viste kunstig DNA seg å være ganske kort. Den inneholder bare 81 basepar. Imidlertid reproduserer og utvikler den seg også.
Replikering av et molekyl oppnådd kunstig skjer takket være polymerasekjedereaksjonen, men så langt skjer dette ikke uavhengig, men gjennom inngripen fra forskere. De tilsetter uavhengig de nødvendige enzymene til nevnte DNA, og plasserer det i et spesielt forberedt flytende medium.
Endelig resultat
Prosessen og det endelige resultatet av DNA-utvikling kan påvirkes av ulike faktorer, for eksempel mutasjoner. Dette gjør det nødvendig å studere stoffprøver slik at analyseresultatet blir pålitelig og pålitelig. Et eksempel er en farskapstest. Men vi kan ikke annet enn å glede oss over at hendelser som mutasjon er sjeldne. Likevel blir prøver av stoff alltid kontrollert på nytt for å få mer nøyaktig informasjon basert på analysen.

Plante-DNA
Takket være høysekvenseringsteknologier (HTS) har det blitt gjort en revolusjon innen genomikk - DNA-ekstraksjon fra planter er også mulig. Å skaffe høykvalitets molekylvekts-DNA fra plantemateriale utgjør selvfølgelig noen vanskeligheter på grunn av det store antallet kopier av mitokondrier og kloroplast-DNA, samt det høye nivået av polysakkarider og fenoliske forbindelser. For å isolere strukturen vi vurderer i dette tilfellet, brukes en rekke metoder.
Hydrogenbinding i DNA
Hydrogenbindingen i DNA-molekylet er ansvarlig for den elektromagnetiske tiltrekningen som skapes mellom et positivt ladet hydrogenatom som er festet til et elektronegativt atom. Denne dipolinteraksjonen oppfyller ikke kriteriet om en kjemisk binding. Men det kan forekomme intermolekylært eller i forskjellige deler av molekylet, dvs. intramolekylært.

Et hydrogenatom fester seg til det elektronegative atomet som er giveren av bindingen. Et elektronegativt atom kan være nitrogen, fluor eller oksygen. Den - gjennom desentralisering - tiltrekker seg elektronskyen fra hydrogenkjernen til seg selv og gjør hydrogenatomet (delvis) positivt ladet. Siden størrelsen på H er liten sammenlignet med andre molekyler og atomer, er ladningen også liten.
DNA-avkoding
Før de dechiffrerer et DNA-molekyl, tar forskerne først et stort antall celler. For det mest nøyaktige og vellykkede arbeidet trengs omtrent en million av dem. Resultatene oppnådd under studien sammenlignes og registreres kontinuerlig. I dag er genomavkoding ikke lenger en sjeldenhet, men en tilgjengelig prosedyre.

Å dechiffrere genomet til en enkelt celle er selvfølgelig en upraktisk øvelse. Dataene innhentet under slike studier er ikke av interesse for forskere. Men det er viktig å forstå at alle eksisterende dekodingsmetoder, til tross for deres kompleksitet, ikke er effektive nok. De vil bare tillate lesing av 40-70% av DNA.
Harvard-professorer kunngjorde imidlertid nylig en metode der 90 % av genomet kan dechiffreres. Teknikken er basert på å tilsette primermolekyler til isolerte celler, ved hjelp av disse starter DNA-replikasjonen. Men selv denne metoden kan ikke betraktes som vellykket; den må fortsatt raffineres før den kan brukes åpent i vitenskapen.
