Molekula e ADN-së ndodhet. ADN dhe gjenet. Vështrim i ri në ADN
Bazat biokimike të trashëgimisë.
Roli gjenetik i acideve nukleike.
Acidet nukleike janë polimere biologjike që gjenden në të gjitha qelizat, nga ato primitive në ato komplekse. Ato u zbuluan për herë të parë nga Johann Friedrich Miescher në 1868 në qeliza të pasura me material bërthamor (leukocite, sperma e salmonit). Termi "acidet nukleike" u propozua në 1889.
Ekzistojnë dy lloje të acideve nukleike: ADN, ARN (ATP - mononukleotid). ADN dhe ARN janë molekula - matrica. ADN-ja përmbahet rreth 6 * 10 -12 g në qelizat somatike: në bërthamë, mitokondri. ARN është pjesë e ribozomeve dhe gjendet në bërthamë dhe në citoplazmë.
Studimi dhe vërtetimi i rolit drejtues të acideve nukleike në transmetimin e informacionit trashëgues u krye në grimcat virale. Virusi i mozaikut të duhanit dihet se është virulent për duhanin dhe delli. Grimca virale përbëhet nga 95% proteina dhe 5% acid nukleik. Kapsidi i proteinës në grimcat virale u ndërrua, por pas disa kohësh proteina në të dy shtamet u shndërrua në formën e mëparshme.
Në bakteriofagët që infektojnë E. coli, proteinat e shtresës së fagut u etiketuan me S radioaktive dhe ADN-ja e fagut u etiketua me P radioaktive. Në një qelizë bakteriale të infektuar me fagun, u formuan grimca fag që përmbanin vetëm P radioaktiv.
Struktura dhe funksionet e molekulave të ADN-së dhe ARN-së.
Acidet nukleike janë biopolimere me strukturë të parregullt, monomerët e të cilëve janë nukleotide. Nukleotidi përbëhet nga mbetjet e tre substancave: acid fosforik, karbohidrate - pentozë, bazë azotike. Nukleotidet e ADN-së përmbajnë karbohidratin deoksiribozë, dhe ARN përmban ribozë. Mbetjet e bazave azotike purine dhe pirimidine që përbëjnë ADN-në janë adenina, guanina, citozina, timina. Molekulat e ARN-së përmbajnë adeninë, guaninë, citozinë dhe uracil.
Nukleotidet janë të lidhura me njëri-tjetrin nëpërmjet mbetjes së acidit fosforik të një nukleotidi dhe karbohidrateve të një tjetri nga një lidhje e fortë kovalente esterike, e quajtur "urë oksigjeni". Lidhja kalon përmes atomit të 5-të të karbonit të një karbohidrati të një nukleotidi në atomin e tretë të karbonit të një karbohidrati të një nukleotidi tjetër. Sekuenca nukleotide përfaqëson strukturën parësore të acideve nukleike. ARN është një zinxhir i vetëm polinukleotid. Struktura e ADN-së është një zinxhir i dyfishtë polinukleotid, i mbështjellë në një spirale.
Struktura dytësore e ADN-së formohet kur shfaqet një varg i dytë i ADN-së, i rregulluar sipas parimit të komplementaritetit në lidhje me të parën. Zinxhiri i dytë është i kundërt me të parin (antiparalel). Bazat azotike shtrihen në një plan pingul me rrafshin e molekulës - kjo i ngjan një shkalle spirale. Kangjellat e kësaj shkalle janë mbetje të acidit fosforik dhe karbohidrate, dhe shkallët janë baza azotike.
Bazat azotike që përbëjnë çdo nukleotid në zinxhirë të drejtuar në mënyrë të kundërt janë të afta të formojnë lidhje hidrogjenore plotësuese me njëra-tjetrën (për shkak të grupeve funksionale ekzistuese në strukturën e secilës bazë azotike). Adenil nukleotidi është plotësues i timinës, guanil ndaj citozinës dhe anasjelltas. Në vetvete, këto lidhje janë të brishta, por një molekulë e ADN-së e "qepur" në mënyrë të përsëritur përgjatë gjithë gjatësisë së saj me lidhje të tilla përfaqëson një lidhje shumë të fortë.
Komplementariteti- kjo është korrespondenca hapësinore-strukturore dhe kimike e bazave azotike me njëra-tjetrën; ato përshtaten me njëra-tjetrën "si një çelës në një bravë".
Një molekulë e ADN-së mund të përmbajë 10 8 ose më shumë nukleotide.
Struktura e molekulës së ADN-së si një spirale e dyfishtë antiparalele u propozua në vitin 1953 nga biologu amerikan James Watson dhe fizikani anglez Francis Crick.
Molekula e ADN-së e çdo organizmi të gjallë në planet përbëhet nga vetëm katër lloje nukleotidesh, të cilat ndryshojnë nga njëri-tjetri në bazat azotike që përmbajnë: adenil, guanil, timinë dhe citozinë. Në atë shkathtësi ADN. Sekuenca e tyre është e ndryshme, dhe numri është i pafund.
Për çdo specie të organizmave të gjallë dhe për secilin organizëm veç e veç, kjo sekuencë është individuale dhe rreptësisht specifike .
Veçori Struktura e ADN-së është se seksionet kimikisht aktive të molekulës - bazat azotike, janë zhytur në qendër të spirales dhe formojnë lidhje plotësuese me njëra-tjetrën, dhe mbetjet e deoksiribozës dhe acidit fosforik janë të vendosura në periferi dhe mbulojnë hyrjen në bazat azotike. - janë kimikisht joaktive. Kjo strukturë mund të ruajë stabilitetin kimik për një kohë të gjatë. Çfarë tjetër nevojitet për të ruajtur informacionin trashëgues? Janë këto veçori të strukturës së ADN-së që përcaktojnë aftësinë e saj për të koduar dhe riprodhuar informacionin gjenetik.
Struktura e fortë e ADN-së është mjaft e vështirë për t'u shkatërruar. Sidoqoftë, kjo ndodh rregullisht në qelizë - gjatë sintezës së ARN-së dhe dyfishimit të vetë molekulës së ADN-së para ndarjes së qelizave.
Dyfishim, replikim i ADN-së fillon me faktin se një enzimë e veçantë - polimeraza e ADN-së - zbërthen spiralen e dyfishtë dhe e ndan atë në fije të veçanta - formohet një pirun reduplikimi. Enzima vepron si një bllokues në një zinxhir. Në çdo zinxhir me një fije - skajet ngjitëse të pirunit të riprodhimit - sintetizohet një zinxhir i ri nga nukleotidet e lira në karioplazmë sipas parimit të komplementaritetit. Në dy molekula të reja të ADN-së, njëra varg mbetet fillesa origjinale e nënës dhe e dyta mbetet fillesa e re bijë. Si rezultat, në vend të një molekule të ADN-së, shfaqen dy molekula me të njëjtën përbërje të saktë nukleotide si ajo origjinale.
Në sistemet e gjalla ne hasim një lloj të ri reagimesh të panjohura në natyrën e pajetë. Ata janë quajtur reaksionet e sintezës së matricës . Sinteza e matricës është si hedhja në një matricë: molekulat e reja sintetizohen në përputhje të saktë me planin e përcaktuar në strukturën e molekulave ekzistuese. Këto reaksione sigurojnë sekuencën e saktë të njësive monomere në polimerët e sintetizuar. Monomerët arrijnë në një vendndodhje specifike në molekulat që shërbejnë si një matricë ku ndodh reaksioni. Nëse reagime të tilla do të ndodhnin si rezultat i përplasjeve të rastësishme të molekulave, ato do të vazhdonin pafundësisht ngadalë. Sinteza e molekulave komplekse bazuar në parimin e shabllonit kryhet shpejt dhe saktë me ndihmën e enzimave. Sinteza e shabllonit qëndron në bazë të reaksioneve më të rëndësishme në sintezën e acideve nukleike dhe proteinave. Roli i matricës në qelizë luhet nga molekulat e acideve nukleike ADN ose ARN. Molekulat monomerike nga të cilat sintetizohet polimeri - nukleotidet ose aminoacidet - në përputhje me parimin e komplementaritetit, vendosen dhe fiksohen në matricë në një rend të përcaktuar rreptësisht. Pastaj njësitë e monomerit lidhen në një zinxhir polimer, dhe polimeri i përfunduar largohet nga matrica. Pas kësaj, matrica është gati për montimin e një molekule të re, saktësisht të njëjtë të polimerit.
Reaksionet e tipit matricë janë një tipar specifik i një qelize të gjallë. Ato janë baza e pronës themelore të të gjitha gjallesave - aftësia për të riprodhuar llojin e tyre.
Funksionet e acideve nukleike– ruajtja dhe transmetimi i informacionit trashëgues. Molekulat e ADN-së kodojnë informacion rreth strukturës parësore të proteinës. Sinteza e molekulave të mARN-së ndodh në matricën e ADN-së. Ky proces quhet "transkriptim". I-ARN në procesin e "përkthimit" zbaton informacionin në formën e një sekuence të aminoacideve në një molekulë proteine.
ADN-ja e secilës qelizë mbart informacion jo vetëm për proteinat strukturore që përcaktojnë formën e qelizës, por edhe për të gjitha proteinat enzimë, proteinat hormonale dhe proteinat e tjera, si dhe strukturën e të gjitha llojeve të ARN-së.
Ndoshta acidet nukleike ofrojnë lloje të ndryshme të memories biologjike - imunologjike, neurologjike, etj., Dhe gjithashtu luajnë një rol të rëndësishëm në rregullimin e proceseve biosintetike.
Informacione të lidhura.
Prova e parë e rolit të ADN-së si bartës i informacionit trashëgues në organizma tërhoqi vëmendje të madhe në studimin e acideve nukleike. Në vitin 1869, F. Miescher izoloi një substancë të veçantë nga bërthamat e qelizave, të cilën ai e quajti nukleinë. Pas 20 vitesh ky emër u zëvendësua me termin acidi nukleik. Në vitin 1924, R. Felgen zhvilloi një metodë për njohjen citologjike të acideve nukleike përmes ngjyrosjes specifike të tyre dhe tregoi se ADN-ja është e lokalizuar në bërthamat e qelizave, dhe ARN në citoplazmë. Në vitin 1936 A.N. Belozersky dhe I.I. Dubrovskaya izoloi ADN-në në formën e saj të pastër nga bërthamat e qelizave bimore. Nga fillimi i viteve 1930. Parimet themelore kimike të strukturës së sheqernave të acidit nukleik u sqaruan dhe në 1953 u krijua një model strukturor i ADN-së.
Njësia bazë strukturore e acideve nukleike është nukleotidi, i cili përbëhet nga tre pjesë kimikisht të ndryshme të lidhura me lidhje kovalente (Fig. 5.2).
Oriz. 5.2. Formulat strukturore: A- nukleotide; b- ADN; V - ARN (shih gjithashtu f. 110)

Oriz. 5.2. Përfundimi. Formulat strukturore: A- nukleotide; 6 - ADN; V- ARN
Pjesa e parë është sheqeri që përmban pesë atome karboni: deoksiriboza në ADN dhe ribozë në ARN.
Pjesa e dytë e nukleotidit, një bazë azotike purine ose pirimidine, është e lidhur në mënyrë kovalente me atomin e parë të karbonit të sheqerit, duke formuar një strukturë të quajtur nukleozid. ADN-ja përmban baza purine - adenina(A) dhe guaninë(D) - dhe bazat pirimidine - timinë(T) dhe citozina(C). Nukleozidet përkatëse quhen deoksiadenozinë, deoksiguanozinë, deoksitimidinë dhe deoksicitidinë. ARN përmban të njëjtat baza purine si ADN-ja, një bazë pirimidine citozina, dhe në vend të timinës përmban uracil(U); nukleozidet përkatëse quhen adenozinë, guanozinë, uridinë dhe citidinë.
Pjesa e tretë e nukleotidit është një grup fosfat, i cili lidh nukleozidet fqinje në një zinxhir polimer përmes lidhjeve fosfodiesterike midis atomit 5-karbon të një sheqeri dhe atomit 3-karbon të një tjetri (Fig. 5.2, b, V). Nukleotidet quhen nukleozide me një ose më shumë grupe fosfate të lidhura me lidhje estere në atomet 3" ose 5 të karbonit të sheqerit. Sinteza e nukleotideve i paraprin sintezës së acideve nukleike; në përputhje me rrethanat, nukleotidet janë produkte të hidrolizës kimike ose enzimatike të acideve nukleike.
Acidet nukleike janë zinxhirë polimerësh shumë të gjatë të përbërë nga mononukleotide të lidhura me lidhje 5- dhe 3'-fosfodiester. Një molekulë e paprekur e ADN-së përmban, në varësi të llojit të organizmit, nga disa mijëra në shumë miliona nukleotide, një molekulë të paprekur ARN - nga 100 në 100 mijë ose më shumë nukleotide.
Rezultatet e analizave të E. Chargaff të përbërjes nukleotide të ADN-së të formave të ndryshme të specieve treguan se raporti molekular i bazave të ndryshme azotike - adenina, guanina, timina, citozina - ndryshon shumë. Rrjedhimisht, u vërtetua se ADN-ja nuk është aspak një polimer monoton i përbërë nga tetranukleotide identike, siç supozohej në vitet '40. shekuj XX, dhe se zotëron plotësisht kompleksitetin e nevojshëm për ruajtjen dhe transmetimin e informacionit trashëgues në formën e një sekuence specifike të bazave nukleotide.
Hulumtimi nga E. Chargaff zbuloi gjithashtu një veçori të natyrshme në të gjitha molekulat e ADN-së: përmbajtja molare e adeninës është e barabartë me përmbajtjen e timinës dhe përmbajtja molare e guaninës është e barabartë me përmbajtjen e citozinës. Këto barazi quhen rregulli i ekuivalencës së Chargaff: [A] = [T], [G] = [C]; numri i purinave është i barabartë me numrin e pirimidinave. Në varësi të llojit, vetëm raporti ([A] + [T])/([G] + [C]) ndryshon (Tabela 5.1).
|
Përbërja e bazave |
Qëndrimi |
Asimetria |
|||||
|
arsye |
|||||||
|
(A + T)/(G + C) |
|||||||
|
Kafshët |
|||||||
|
breshkë |
|||||||
|
gaforre deti |
|||||||
|
Iriq Deti |
|||||||
|
Bimët, kërpudhat |
|||||||
|
embrion gruri |
|||||||
|
Kërpudha Aspergillus niger |
|||||||
|
Bakteret |
|||||||
|
Escherichia coli |
|||||||
|
Staphylococcus aureus |
|||||||
|
Clostridium perfringens |
|||||||
|
Brucela abortus |
|||||||
|
Sarcina lutea |
|||||||
|
Bakteriofagët |
|||||||
|
FH 174 (formë virale) |
|||||||
|
FH 174 (formë replikuese) |
|||||||
Raporti i bazave quhet koeficienti nukleotid(specie) specifika. Zbulimi i Chargaff formuloi një veçori të rëndësishme strukturore të ADN-së, e cila më vonë u pasqyrua në modelin strukturor të ADN-së nga J. Watson dhe F. Crick (1953), të cilët në fakt treguan se rregullat e Chargaff nuk vendosin asnjë kufizim në numrin e mundshëm të kombinimeve të sekuenca të ndryshme bazë të afta për të formuar molekulat e ADN-së.
Koncepti i specifikës së nukleotideve formoi bazën e një dege të re të biologjisë - sistematika e gjeneve, i cili vepron duke krahasuar përbërjen dhe strukturën e acideve nukleike për të ndërtuar një sistem natyror organizmash.
Sipas modelit Watson-Crick, një molekulë e ADN-së përbëhet nga dy zinxhirë polinukleotide (vargje, fije), të lidhura me njëri-tjetrin duke përdorur lidhje hidrogjenore tërthore midis bazave azotike sipas parimit plotësues (adeni i një zinxhiri është i lidhur me dy lidhje hidrogjeni me timina e zinxhirit të kundërt, dhe guanina dhe citozina e vargjeve të ndryshme janë të lidhura me njëra-tjetrën me tre lidhje hidrogjenore). Në këtë rast, dy vargje polinukleotide të një molekule janë antiparalele, d.m.th., përballë skajit 3" të njërit zinxhir është fundi 5" i zinxhirit tjetër dhe anasjelltas (Fig. 5.3). Megjithatë, duhet mbajtur parasysh të dhënat moderne se materiali gjenetik i disa viruseve përfaqësohet nga molekula të ADN-së me një fije (një-vargësh). Bazuar në analizën e difraksionit me rreze X të ADN-së, J. Watson dhe F. Crick arritën gjithashtu në përfundimin se molekula e saj me dy fije ka një strukturë dytësore në formën e një spiraleje të përdredhur në drejtim nga e majta në të djathtë, e cila më vonë u quajt 5. -formë (Fig. 5.4). Deri më sot, është vërtetuar se përveç formës më të zakonshme 5, është e mundur të zbulohen seksione të ADN-së që kanë një konfigurim të ndryshëm - si të djathtë (forma A, C), dhe i përdredhur nga e djathta në të majtë (i përdredhur majtas, ose në formë Z) (Fig. 5.4). Ekzistojnë disa dallime midis këtyre formave të strukturës sekondare të ADN-së (Tabela 5.2). Për shembull, distanca midis dy çifteve ngjitur të bazave azotike në një spirale me dy fije, e shprehur në nanometra (nm), karakterizohet nga vlera të ndryshme për formën 5 dhe Z-formën (0,34 dhe 0,38 nm, respektivisht). . Në Fig. Figura 5.5 tregon modelet moderne tre-dimensionale të formave të ADN-së "me dorën e majtë" dhe "të djathtën".

Oriz. 5.3. paraqitje skematike e strukturës primare të një fragmenti të një molekule të ADN-së me dy fije: A - adeninë; G - guaninë; T - timinë; C - citozinë

Oriz. 5.4.

Tabela 5.2
Vetitë e formave të ndryshme të helikave të dyfishta të ADN-së
Molekulat e ARN-së, në varësi të karakteristikave të tyre strukturore dhe funksionale, ndahen në disa lloje: ARN lajmëtare (mARN, ose mARN), ARN ribozomale (rARN), ARN transferuese (tARN), ARN e vogël bërthamore (snARN), etj. Molekulat e ADN-së, ARN-së janë gjithmonë njëvargëshe (një-vargëshe). Sidoqoftë, ato mund të formojnë konfigurime më komplekse (sekondare) për shkak të lidhjes plotësuese të seksioneve individuale të një zinxhiri të tillë bazuar në ndërveprimin e bazave plotësuese azotike (A-U dhe G-C). Si shembull, merrni parasysh konfigurimin e gjetheve të tërfilit për molekulën e ARN-së së transferimit të fenilalaninës (Figura 5.6).

Oriz. 5.6.
Në vitin 1953, D. Watson dhe F. Crick propozuan një model të strukturës së ADN-së, i cili bazohej në postulatet e mëposhtme:
- 1. ADN-ja është një polimer i përbërë nga nukleotide të lidhura me lidhje fosfodiesterike 3" dhe 5".
- 2. Përbërja e nukleotideve të ADN-së i bindet rregullave të Chargaff-it.
- 3. Molekula e ADN-së ka një strukturë spirale të dyfishtë, që të kujton një shkallë spirale, siç dëshmohet nga modelet e difraksionit me rreze X të vargjeve të ADN-së të marra fillimisht nga M. Wilkins dhe R. Franklin.
- 4. Struktura e polimerit, siç tregohet nga titrimi acid-bazë i ADN-së vendase (natyrore), stabilizohet nga lidhjet hidrogjenore. Titrimi dhe ngrohja e ADN-së vendase shkakton një ndryshim të dukshëm në vetitë e saj fizike, në veçanti viskozitetin, duke e shndërruar atë në një formë të denatyruar dhe lidhjet kovalente nuk shkatërrohen.
Acidet nukleike janë substanca me molekulare të lartë të përbëra nga mononukleotide, të cilat janë të lidhura me njëri-tjetrin në një zinxhir polimer duke përdorur lidhje fosfodiesterike 3", 5" dhe paketohen në qeliza në një mënyrë të caktuar.
Acidet nukleike janë biopolimere të dy llojeve: acidi ribonukleik (ARN) dhe acidi deoksiribonukleik (ADN). Çdo biopolimer përbëhet nga nukleotide që ndryshojnë në mbetjen e karbohidrateve (ribozë, deoksiribozë) dhe një nga bazat azotike (uracil, timinë). Sipas këtyre dallimeve, acidet nukleike morën emrin e tyre.
Struktura e acidit deoksiribonukleik
Acidet nukleike kanë një strukturë parësore, dytësore dhe terciare.
Struktura primare e ADN-së
Struktura primare e ADN-së është një zinxhir polinukleotid linear në të cilin mononukleotidet janë të lidhura me lidhje fosfodiesterike 3", 5". Materiali fillestar për montimin e një zinxhiri të acidit nukleik në një qelizë është nukleozidi 5"-trifosfat, i cili, si rezultat i heqjes së mbetjeve të acidit fosforik β dhe γ, është i aftë të bashkojë atomin e karbonit 3" të një nukleozidi tjetër. . Kështu, atomi i karbonit 3" i një dezoksiriboze është i lidhur në mënyrë kovalente me atomin e karbonit 5" të një deoksiriboze tjetër përmes një mbetjeje të vetme të acidit fosforik dhe formon një zinxhir linear polinukleotid të acidit nukleik. Prandaj emri: lidhje fosfodiesterike 3", 5". Bazat azotike nuk marrin pjesë në lidhjen e nukleotideve të një zinxhiri (Fig. 1.).
Një lidhje e tillë, ndërmjet mbetjes së molekulës së acidit fosforik të një nukleotidi dhe karbohidratit të një tjetri, çon në formimin e një skeleti pentozë-fosfat të molekulës polinukleotide, mbi të cilin bazat azotike ngjiten anash njëra pas tjetrës. Sekuenca e tyre e rregullimit në zinxhirët e molekulave të acidit nukleik është rreptësisht specifike për qelizat e organizmave të ndryshëm, d.m.th. ka karakter specifik (rregulli i Chargaff-it).
Një zinxhir linear i ADN-së, gjatësia e të cilit varet nga numri i nukleotideve të përfshira në zinxhir, ka dy skaje: njëri quhet fundi 3" dhe përmban një hidroksil të lirë, dhe tjetri quhet fundi 5" dhe përmban një fosfor. mbetje acide. Qarku është polar dhe mund të ketë drejtim 5"->3" dhe 3"->5". Përjashtim është ADN-ja rrethore.
"Teksti" gjenetik i ADN-së përbëhet nga "fjalë" kodike - treshe të nukleotideve të quajtura kodone. Seksionet e ADN-së që përmbajnë informacion në lidhje me strukturën parësore të të gjitha llojeve të ARN quhen gjene strukturore.
Zinxhirët e ADN-së polinukleotidike arrijnë madhësi gjigante, kështu që ato paketohen në një mënyrë të caktuar në qelizë.
Gjatë studimit të përbërjes së ADN-së, Chargaff (1949) vendosi modele të rëndësishme në lidhje me përmbajtjen e bazave individuale të ADN-së. Ato ndihmuan në zbulimin e strukturës dytësore të ADN-së. Këto modele quhen rregullat e Chargaff-it. Rregullat e Chargaff
Këto rregulla tregojnë se gjatë ndërtimit të ADN-së, duhet të respektohet një korrespondencë (çiftim) mjaft i rreptë jo i bazave purine dhe pirimidine në përgjithësi, por veçanërisht e timinës me adeninën dhe citozinës me guaninën. Bazuar në këto rregulla, në vitin 1953, Watson dhe Crick propozuan një model të strukturës dytësore të ADN-së, të quajtur spirale e dyfishtë (Fig.). |

Struktura dytësore e ADN-së
Struktura dytësore e ADN-së është një spirale e dyfishtë, modeli i së cilës u propozua nga D. Watson dhe F. Crick në 1953.
Parakushtet për krijimin e një modeli të ADN-së
Si rezultat i analizave fillestare, besohej se ADN-ja e çdo origjine përmban të katër nukleotidet në sasi molare të barabarta. Megjithatë, në vitet 1940, E. Chargaff dhe kolegët e tij, si rezultat i analizës së ADN-së së izoluar nga një sërë organizmash, treguan qartë se ato përmbanin baza azotike në raporte të ndryshme sasiore. Chargaff zbuloi se megjithëse këto raporte janë të njëjta për ADN-në nga të gjitha qelizat e të njëjtit lloj organizmi, ADN-ja nga specie të ndryshme mund të ndryshojë dukshëm në përmbajtjen e nukleotideve të caktuara. Kjo sugjeroi që ndryshimet në raportin e bazave azotike mund të shoqërohen me një lloj kodi biologjik. Megjithëse raporti i bazave individuale purine dhe pirimidine në mostra të ndryshme të ADN-së doli të ishte i ndryshëm, kur krahasoheshin rezultatet e testit, u shfaq një model i caktuar: në të gjitha mostrat, numri i përgjithshëm i purinave ishte i barabartë me numrin total të pirimidinave (A + G = T + C), sasia e adeninës ishte e barabartë me sasinë e timinës (A = T), dhe sasia e guaninës është sasia e citozinës (G = C). ADN-ja e izoluar nga qelizat e gjitarëve ishte përgjithësisht më e pasur me adeninë dhe timinë dhe relativisht më e varfër me guaninë dhe citozinë, ndërsa ADN-ja nga bakteret ishte më e pasur me guaninë dhe citozinë dhe relativisht më e varfër në adeninë dhe timinë. Këto të dhëna përbënin një pjesë të rëndësishme të materialit faktik mbi bazën e të cilit më vonë u ndërtua modeli Watson-Crick i strukturës së ADN-së.
Një tjetër tregues i rëndësishëm indirekt i strukturës së mundshme të ADN-së u dha nga të dhënat e L. Pauling mbi strukturën e molekulave të proteinave. Pauling tregoi se disa konfigurime të ndryshme të qëndrueshme të zinxhirit të aminoacideve në një molekulë proteine janë të mundshme. Një konfigurim i zakonshëm i zinxhirit peptid, α-spiralja, është një strukturë spirale e rregullt. Me këtë strukturë, formimi i lidhjeve hidrogjenore midis aminoacideve të vendosura në kthesat ngjitur të zinxhirit është i mundur. Pauling përshkroi konfigurimin α-spiral të zinxhirit polipeptid në 1950 dhe sugjeroi që molekulat e ADN-së ndoshta kanë një strukturë spirale të mbajtur në vend nga lidhjet hidrogjenore.
Sidoqoftë, informacioni më i vlefshëm në lidhje me strukturën e molekulës së ADN-së u dha nga rezultatet e analizës së difraksionit me rreze X. Rrezet X që kalojnë nëpër një kristal të ADN-së i nënshtrohen difraksionit, domethënë ato devijohen në drejtime të caktuara. Shkalla dhe natyra e devijimit të rrezeve varet nga struktura e vetë molekulave. Një model difraksioni me rreze X (Fig. 3) i jep syrit me përvojë një sërë indikacionesh indirekte në lidhje me strukturën e molekulave të substancës në studim. Analiza e modeleve të difraksionit me rreze X të ADN-së çoi në përfundimin se bazat azotike (të cilat kanë një formë të sheshtë) janë të rregulluara si një pirg pllakash. Modelet e difraksionit me rreze X zbuluan tre periudha kryesore në strukturën e ADN-së kristalore: 0.34, 2 dhe 3.4 nm.

Modeli i ADN-së Watson-Crick
Bazuar në të dhënat analitike të Chargaff-it, modelet e rrezeve X të Wilkins dhe kërkimin e kimistëve që dhanë informacione rreth distancave të sakta midis atomeve në një molekulë, këndeve midis lidhjeve të një atomi të caktuar dhe madhësisë së atomeve, Watson dhe Crick filloi të ndërtojë modele fizike të përbërësve individualë të molekulës së ADN-së në një shkallë të caktuar dhe t'i "rregullojë" ato me njëri-tjetrin në atë mënyrë që sistemi që rezulton të korrespondojë me të dhëna të ndryshme eksperimentale. [shfaqje] .
Dihej edhe më herët se nukleotidet fqinje në një zinxhir të ADN-së janë të lidhura me ura fosfodiesterike, duke lidhur atomin 5"-deoksiriboz të karbonit të një nukleotidi me atomin 3"-deoksiriboz të karbonit të nukleotidit tjetër. Watson dhe Crick nuk kishin dyshim se periudha prej 0.34 nm korrespondon me distancën midis nukleotideve të njëpasnjëshme në zinxhirin e ADN-së. Më tej, mund të supozohet se periudha prej 2 nm korrespondon me trashësinë e zinxhirit. Dhe për të shpjeguar se me çfarë strukture reale korrespondon periudha prej 3.4 nm, Watson dhe Crick, si dhe Pauling më herët, sugjeruan që zinxhiri është i përdredhur në formën e një spirale (ose, më saktë, formon një vijë spirale, pasi një spirale në kuptimin e ngushtë të kësaj fjale fitohen kur mbështjelljet formojnë një sipërfaqe konike dhe jo cilindrike në hapësirë). Pastaj një periudhë prej 3.4 nm do të korrespondojë me distancën midis kthesave të njëpasnjëshme të kësaj spiraleje. Një spirale e tillë mund të jetë shumë e dendur ose disi e shtrirë, domethënë kthesat e saj mund të jenë të sheshta ose të pjerrëta. Meqenëse periudha prej 3,4 nm është saktësisht 10 herë distanca midis nukleotideve të njëpasnjëshme (0,34 nm), është e qartë se çdo kthesë e plotë e spirales përmban 10 nukleotide. Nga këto të dhëna, Watson dhe Crick ishin në gjendje të llogarisin densitetin e një zinxhiri polinukleotid të përdredhur në një spirale me një diametër prej 2 nm, me një distancë midis kthesave prej 3.4 nm. Doli se një zinxhir i tillë do të kishte një densitet sa gjysma e densitetit aktual të ADN-së, e cila dihej tashmë. Më duhej të supozoja se molekula e ADN-së përbëhet nga dy zinxhirë - se është një spirale e dyfishtë nukleotidesh.
Detyra tjetër ishte, natyrisht, të qartësonte marrëdhëniet hapësinore midis dy zinxhirëve që formojnë spiralen e dyfishtë. Duke provuar një sërë opsionesh për rregullimin e zinxhirëve në modelin e tyre fizik, Watson dhe Crick zbuluan se të gjitha të dhënat e disponueshme përputheshin më së miri me opsionin në të cilin dy spirale polinukleotide shkojnë në drejtime të kundërta; në këtë rast, zinxhirët e përbërë nga mbetje sheqeri dhe fosfati formojnë sipërfaqen e spirales së dyfishtë, dhe purinat dhe pirimidinat janë të vendosura brenda. Bazat e vendosura përballë njëra-tjetrës, që i përkasin dy zinxhirëve, lidhen në çift me lidhje hidrogjenore; Janë këto lidhje hidrogjenore që mbajnë zinxhirët së bashku, duke fiksuar kështu konfigurimin e përgjithshëm të molekulës.
Spiralja e dyfishtë e ADN-së mund të imagjinohet si një shkallë litari që është e përdredhur në mënyrë spirale, në mënyrë që shkallët e saj të mbeten horizontale. Pastaj dy litarët gjatësorë do të korrespondojnë me zinxhirët e mbetjeve të sheqerit dhe fosfatit, dhe shufrat do të korrespondojnë me çifte bazash azotike të lidhura me lidhje hidrogjeni.
Si rezultat i studimit të mëtejshëm të modeleve të mundshme, Watson dhe Crick arritën në përfundimin se çdo "shirit" duhet të përbëhet nga një purinë dhe një pirimidinë; në një periudhë prej 2 nm (që korrespondon me diametrin e spirales së dyfishtë), nuk do të kishte hapësirë të mjaftueshme për dy purina dhe dy pirimidinat nuk mund të ishin aq afër njëra-tjetrës për të formuar lidhjet e duhura hidrogjenore. Një studim i thelluar i modelit të detajuar tregoi se adenina dhe citozina, ndërsa formonin një kombinim të një madhësie të përshtatshme, ende nuk mund të pozicionoheshin në atë mënyrë që të krijoheshin lidhje hidrogjeni midis tyre. Raportime të ngjashme detyruan përjashtimin e kombinimit guaninë - timinë, ndërsa kombinimet adeninë - timinë dhe guaninë - citozinë rezultuan mjaft të pranueshme. Natyra e lidhjeve hidrogjenore është e tillë që adenina formon një çift me timinën dhe guanina me citozinën. Kjo ide e çiftimit të bazave specifike bëri të mundur shpjegimin e "rregullit Chargaff", sipas të cilit në çdo molekulë të ADN-së sasia e adeninës është gjithmonë e barabartë me përmbajtjen e timinës, dhe sasia e guaninës është gjithmonë e barabartë me sasinë. të citozinës. Dy lidhje hidrogjenore formohen midis adeninës dhe timinës, dhe tre midis guaninës dhe citozinës. Për shkak të kësaj specifike, formimi i lidhjeve hidrogjenore kundër çdo adenine në një zinxhir shkakton formimin e timinës në tjetrin; në të njëjtën mënyrë, vetëm citozina mund të jetë përballë çdo guanine. Kështu, zinxhirët janë plotësues me njëri-tjetrin, domethënë sekuenca e nukleotideve në një zinxhir përcakton në mënyrë unike sekuencën e tyre në tjetrën. Dy zinxhirët shkojnë në drejtime të kundërta dhe grupet e tyre fundore të fosfatit janë në skajet e kundërta të spirales së dyfishtë.
Si rezultat i hulumtimit të tyre, në vitin 1953 Watson dhe Crick propozuan një model të strukturës së molekulës së ADN-së (Fig. 3), i cili mbetet i rëndësishëm deri në ditët e sotme. Sipas modelit, molekula e ADN-së përbëhet nga dy zinxhirë polinukleotidikë plotësues. Çdo varg i ADN-së është një polinukleotid i përbërë nga disa dhjetëra mijëra nukleotide. Në të, nukleotidet fqinje formojnë një shtyllë të rregullt pentozë-fosfat për shkak të lidhjes së një mbetje të acidit fosforik dhe deoksiribozës nga një lidhje e fortë kovalente. Bazat azotike të njërit zinxhir polinukleotid janë të rregulluara në një rend të përcaktuar rreptësisht përballë bazave azotike të tjetrit. Alternimi i bazave azotike në një zinxhir polinukleotid është i parregullt.
Rregullimi i bazave azotike në zinxhirin e ADN-së është plotësues (nga greqishtja "komplement" - shtesë), d.m.th. Timina (T) është gjithmonë kundër adeninës (A), dhe vetëm citozina (C) është kundër guaninës (G). Kjo shpjegohet me faktin se A dhe T, si dhe G dhe C, korrespondojnë rreptësisht me njëra-tjetrën, d.m.th. plotësojnë njëra-tjetrën. Kjo korrespondencë përcaktohet nga struktura kimike e bazave, e cila lejon formimin e lidhjeve hidrogjenore në çiftin purine dhe pirimidine. Ka dy lidhje midis A dhe T, dhe tre midis G dhe C. Këto lidhje sigurojnë stabilizim të pjesshëm të molekulës së ADN-së në hapësirë. Qëndrueshmëria e spirales së dyfishtë është drejtpërdrejt proporcionale me numrin e lidhjeve G≡C, të cilat janë më të qëndrueshme në krahasim me lidhjet A=T.
Sekuenca e njohur e renditjes së nukleotideve në një zinxhir të ADN-së bën të mundur, sipas parimit të komplementaritetit, vendosjen e nukleotideve të një zinxhiri tjetër.
Për më tepër, është vërtetuar se bazat azotike që kanë një strukturë aromatike në një zgjidhje ujore janë të vendosura njëra mbi tjetrën, duke formuar, si të thuash, një pirg monedhash. Ky proces i formimit të pirgjeve të molekulave organike quhet grumbullim. Zinxhirët polinukleotid të molekulës së ADN-së të modelit Watson-Crick në shqyrtim kanë një gjendje të ngjashme fiziko-kimike, bazat e tyre azotike janë rregulluar në formën e një pirg monedhash, midis planeve të të cilave lindin ndërveprimet van der Waals (ndërveprimet e grumbullimit).
Lidhjet e hidrogjenit midis bazave plotësuese (horizontale) dhe ndërveprimet e grumbullimit midis planeve të bazave në një zinxhir polinukleotid për shkak të forcave van der Waals (vertikalisht) i sigurojnë molekulës së ADN-së stabilizim shtesë në hapësirë.
Kockat kurrizore të fosfatit të sheqerit të të dy zinxhirëve janë të kthyera nga jashtë, dhe bazat janë të kthyera nga brenda, drejt njëra-tjetrës. Drejtimi i zinxhirëve në ADN është antiparalel (njëri prej tyre ka drejtim 5"->3", tjetri - 3"->5", pra skaji 3" i njërit zinxhir ndodhet përballë skajit 5" të tjetri.). Zinxhirët formojnë spirale të djathta me një bosht të përbashkët. Një kthesë e spirales është 10 nukleotide, madhësia e kthesës është 3.4 nm, lartësia e secilit nukleotid është 0.34 nm, diametri i spirales është 2.0 nm. Si rezultat i rrotullimit të një vargu rreth tjetrit, formohet një brazdë e madhe (rreth 20 Å në diametër) dhe një brazdë e vogël (rreth 12 A në diametër) të spirales së dyfishtë të ADN-së. Kjo formë e spirales së dyfishtë Watson-Crick u quajt më vonë forma B. Në qeliza, ADN-ja zakonisht ekziston në formën B, e cila është më e qëndrueshme.
Funksionet e ADN-së
Modeli i propozuar shpjegoi shumë veti biologjike të acidit deoksiribonukleik, duke përfshirë ruajtjen e informacionit gjenetik dhe diversitetin e gjeneve të ofruara nga një shumëllojshmëri e gjerë kombinimesh sekuenciale të 4 nukleotideve dhe faktin e ekzistencës së një kodi gjenetik, aftësinë për t'u vetë-riprodhuar. dhe transmetojnë informacionin gjenetik të siguruar nga procesi i replikimit, dhe zbatimi i informacionit gjenetik në formën e proteinave, si dhe çdo përbërje tjetër të formuar me ndihmën e proteinave enzimë.
Funksionet themelore të ADN-së.
- ADN-ja është bartës i informacionit gjenetik, i cili sigurohet nga fakti i ekzistencës së një kodi gjenetik.
- Riprodhimi dhe transmetimi i informacionit gjenetik nëpër gjenerata të qelizave dhe organizmave. Ky funksion ofrohet nga procesi i replikimit.
- Zbatimi i informacionit gjenetik në formën e proteinave, si dhe çdo përbërje tjetër të formuar me ndihmën e proteinave enzimë. Ky funksion ofrohet nga proceset e transkriptimit dhe të përkthimit.

Format e organizimit të ADN-së me dy vargje
ADN-ja mund të formojë disa lloje spiralesh të dyfishta (Fig. 4). Aktualisht, gjashtë forma janë tashmë të njohura (nga forma A në E dhe Z).
Format strukturore të ADN-së, siç vendosi Rosalind Franklin, varen nga ngopja e molekulës së acidit nukleik me ujë. Në studimet e fibrave të ADN-së duke përdorur analizën e difraksionit me rreze X, u tregua se modeli i rrezeve X varet rrënjësisht nga lagështia relative në cilën shkallë të ngopjes me ujë të kësaj fije zhvillohet eksperimenti. Nëse fibra ishte mjaftueshëm e ngopur me ujë, atëherë merret një radiografi. Kur u tha, u shfaq një model krejtësisht i ndryshëm i rrezeve X, shumë i ndryshëm nga modeli i rrezeve X të fibrave me lagështi të lartë.
Molekula e ADN-së me lagështi të lartë quhet forma B. Në kushte fiziologjike (përqendrimi i ulët i kripës, shkalla e lartë e hidratimit), lloji strukturor dominues i ADN-së është forma B (forma kryesore e ADN-së me dy vargje - modeli Watson-Crick). Lartësia e spirales së një molekule të tillë është 3.4 nm. Ka 10 çifte plotësuese për kthesë në formën e pirgjeve të përdredhur të "monedhave" - bazave azotike. Pirgjet mbahen së bashku nga lidhjet hidrogjenore midis dy "monedhave" të kundërta të pirgjeve dhe "plagosen" nga dy shirita të shtyllës kurrizore fosfodiesteri të përdredhur në një spirale të djathtë. Planet e bazave azotike janë pingul me boshtin e spirales. Çiftet plotësuese ngjitur rrotullohen në lidhje me njëri-tjetrin me 36°. Diametri i spirales është 20Å, me nukleotidin purine që zë 12Å dhe nukleotidin pirimidin 8Å.
Molekula e ADN-së me lagështi më të ulët quhet forma A. Forma A formohet në kushtet e hidratimit më pak të lartë dhe në një përmbajtje më të lartë të joneve Na + ose K +. Ky konformacion spirale më i gjerë djathtas ka 11 çifte bazash për rrotullim. Rrafshët e bazave azotike kanë një prirje më të fortë ndaj boshtit të spirales; ato devijojnë nga normalja në boshtin e spirales me 20°. Kjo nënkupton praninë e një zbrazëtie të brendshme me një diametër prej 5Å. Distanca midis nukleotideve ngjitur është 0.23 nm, gjatësia e kthesës është 2.5 nm dhe diametri i spirales është 2.3 nm.
Forma A e ADN-së fillimisht mendohej të ishte më pak e rëndësishme. Sidoqoftë, më vonë u bë e qartë se forma A e ADN-së, ashtu si forma B, ka një rëndësi të madhe biologjike. Spiralja ARN-ADN në kompleksin shabllon-primer ka formën A, si dhe strukturat spirale ARN-ARN dhe ARN (grupi 2'-hidroksil i ribozës parandalon molekulat e ARN-së të formojnë formën B). Forma A e ADN-së gjendet në spore. Është vërtetuar se forma A e ADN-së është 10 herë më rezistente ndaj rrezeve UV sesa forma B.
Forma A dhe forma B quhen forma kanonike të ADN-së.
Format C-E gjithashtu me dorën e djathtë, formimi i tyre mund të vërehet vetëm në eksperimente të veçanta, dhe, me sa duket, ato nuk ekzistojnë in vivo. Forma C e ADN-së ka një strukturë të ngjashme me ADN-në B. Numri i çifteve të bazave për kthesë është 9,33, gjatësia e rrotullimit të spirales është 3,1 nm. Çiftet e bazave janë të prirura në një kënd prej 8 gradë në krahasim me pozicionin pingul me boshtin. Vrazda janë të ngjashme në madhësi me brazda të B-ADN-së. Në këtë rast, brazda kryesore është disi më e cekët, dhe brazda e vogël është më e thellë. Polnukleotidet natyrale dhe sintetike të ADN-së mund të shndërrohen në formën C.
| Tabela 1. Karakteristikat e disa llojeve të strukturave të ADN-së | |||
| Lloji spirale | A | B | Z |
| Katrani spirale | 0,32 nm | 3,38 nm | 4,46 nm |
| Përdredhje spirale | E drejta | E drejta | Majtas |
| Numri i çifteve të bazave për kthesë | 11 | 10 | 12 |
| Distanca midis planeve bazë | 0,256 nm | 0,338 nm | 0,371 nm |
| Konformacioni i lidhjes glikozidike | anti | anti | anti-C mëkat-G |
| Konformimi i unazës së furanozës | C3"-endo | C2"-endo | C3"-endo-G C2"-endo-C |
| Gjerësia e brazdës, e vogël/e madhe | 1,11/0,22 nm | 0,57/1,17 nm | 0,2/0,88 nm |
| Thellësia e brazdës, e vogël/e madhe | 0,26/1,30 nm | 0,82/0,85 nm | 1,38/0,37 nm |
| Diametri spirale | 2.3 nm | 2,0 nm | 1.8 nm |
Elementet strukturore të ADN-së
(Strukturat jo-kanonike të ADN-së)
Elementet strukturore të ADN-së përfshijnë struktura të pazakonta të kufizuara nga disa sekuenca të veçanta:
|

ADN në formë Z u zbulua në vitin 1979 gjatë studimit të heksanukleotidit d(CG)3 -. Ajo u zbulua nga profesori i MIT Alexander Rich dhe kolegët e tij. Forma Z është bërë një nga elementët strukturorë më të rëndësishëm të ADN-së për shkak të faktit se formimi i saj është vërejtur në rajonet e ADN-së ku purinat alternojnë me pirimidinat (për shembull, 5'-GCGCGC-3'), ose në përsëritje 5 '-CGCGCG-3' që përmban citozinë të metiluar. Një kusht thelbësor për formimin dhe stabilizimin e Z-ADN ishte prania e nukleotideve purine në të në konformacionin syn, të alternuara me bazat pirimidine në konformacionin anti.
Molekulat natyrore të ADN-së ekzistojnë kryesisht në formën B-djathtas nëse nuk përmbajnë sekuenca si (CG)n. Megjithatë, nëse sekuenca të tilla janë pjesë e ADN-së, atëherë këto seksione, kur forca jonike e tretësirës ose kationeve që neutralizojnë ngarkesën negative në kornizën e fosfodiesterit ndryshon, këto seksione mund të shndërrohen në formën Z, ndërsa seksionet e tjera të ADN-së në zinxhiri mbetet në formën klasike B. Mundësia e një tranzicioni të tillë tregon se dy vargjet në spiralen e dyfishtë të ADN-së janë në një gjendje dinamike dhe mund të zbërthehen në lidhje me njëra-tjetrën, duke lëvizur nga forma e djathtë në atë të majtë dhe anasjelltas. Pasojat biologjike të një qëndrueshmërie të tillë, e cila lejon transformimet konformative të strukturës së ADN-së, nuk janë kuptuar ende plotësisht. Besohet se seksionet e Z-ADN-së luajnë një rol të caktuar në rregullimin e shprehjes së gjeneve të caktuara dhe marrin pjesë në rikombinimin gjenetik.
Forma Z e ADN-së është një spirale e dyfishtë me dorën e majtë, në të cilën shtylla kurrizore e fosfodiesterit ndodhet në një model zigzag përgjatë boshtit të molekulës. Prandaj emri i molekulës (zigzag)-DNK. Z-ADN është ADN-ja më pak e përdredhur (12 çifte bazash për kthesë) dhe ADN-ja më e hollë e njohur në natyrë. Distanca midis nukleotideve ngjitur është 0.38 nm, gjatësia e kthesës është 4.56 nm dhe diametri i Z-ADN është 1.8 nm. Për më tepër, pamja e kësaj molekule të ADN-së dallohet nga prania e një brazdë të vetme.
Forma Z e ADN-së është gjetur në qelizat prokariote dhe eukariote. Tani janë marrë antitrupa që mund të dallojnë formën Z nga forma B e ADN-së. Këto antitrupa lidhen me rajone të caktuara të kromozomeve gjigante të qelizave të gjëndrave të pështymës të Drosophila (Dr. melanogaster). Reagimi i lidhjes është i lehtë për t'u monitoruar për shkak të strukturës së pazakontë të këtyre kromozomeve, në të cilat rajonet (disqet) më të dendura kontrastojnë me rajonet më pak të dendura (ndërdisqet). Regjionet Z-ADN janë të vendosura në ndërdisqe. Nga kjo rrjedh se forma Z ekziston në të vërtetë në kushte natyrore, megjithëse madhësitë e seksioneve individuale të formës Z janë ende të panjohura.

(invertorët) janë sekuencat bazë më të famshme dhe më të shpeshta në ADN. Një palindrom është një fjalë ose frazë që lexon njësoj nga e majta në të djathtë dhe anasjelltas. Shembuj të fjalëve ose frazave të tilla janë: KALIZA, KOZAK, PËRMBYTJA DHE TRENDALI I RËNË NË KUPRA E AZORIT. Kur aplikohet në seksionet e ADN-së, ky term (palindrom) nënkupton të njëjtin alternim të nukleotideve përgjatë zinxhirit nga e djathta në të majtë dhe nga e majta në të djathtë (si shkronjat në fjalën "kasolle", etj.).
Një palindrom karakterizohet nga prania e përsëritjeve të përmbysura të sekuencave bazë që kanë simetri të rendit të dytë në lidhje me dy vargje të ADN-së. Sekuenca të tilla, për arsye të qarta, janë vetë-plotësuese dhe kanë tendencë të formojnë struktura me kapëse flokësh ose kryq (Fig.). Fijet e flokëve ndihmojnë proteinat rregullatore të njohin se ku kopjohet teksti gjenetik i ADN-së së kromozomeve.
Kur një përsëritje e përmbysur është e pranishme në të njëjtën varg të ADN-së, sekuenca quhet përsëritje pasqyre. Përsëritjet e pasqyrës nuk kanë veti të vetë-plotësimit dhe, për rrjedhojë, nuk janë të afta të formojnë strukturë flokësh ose kryq. Sekuencat e këtij lloji gjenden pothuajse në të gjitha molekulat e mëdha të ADN-së dhe mund të variojnë nga vetëm disa çifte bazash deri në disa mijëra çifte bazash.
Prania e palindromave në formën e strukturave kryqore në qelizat eukariotike nuk është vërtetuar, megjithëse një numër i caktuar strukturash kryqëzore janë zbuluar in vivo në qelizat E. coli. Prania e sekuencave vetë-plotësuese në ARN ose në ADN me një fije floku është arsyeja kryesore për palosjen e zinxhirit të acidit nukleik në tretësirë në një strukturë të caktuar hapësinore, e karakterizuar nga formimi i shumë "flokeve".

ADN e formës Hështë një spirale e formuar nga tre vargje të ADN-së - një spirale e trefishtë e ADN-së. Është një kompleks i një spirale të dyfishtë Watson-Crick me një varg të tretë të ADN-së me një fije, e cila përshtatet në brazdë të saj kryesore, duke formuar një të ashtuquajtur çift Hoogsteen.
Formimi i një trefishi të tillë ndodh si rezultat i palosjes së një spirale të dyfishtë të ADN-së në atë mënyrë që gjysma e seksionit të saj të mbetet në formën e një spirale të dyfishtë dhe gjysma tjetër është e ndarë. Në këtë rast, një nga helikat e shkëputura formon një strukturë të re me gjysmën e parë të spirales së dyfishtë - një spirale të trefishtë, dhe e dyta rezulton të jetë e pastrukturuar, në formën e një seksioni me një fije floku. Një tipar i këtij tranzicioni strukturor është varësia e tij e mprehtë nga pH e mediumit, protonet e të cilit stabilizojnë strukturën e re. Për shkak të kësaj veçorie, struktura e re u quajt forma H e ADN-së, formimi i së cilës u zbulua në plazmidet e mbimbështjella që përmbajnë rajone homopurine-homopirimidine, të cilat janë një përsëritje pasqyre.
Në studime të mëtejshme, u vërtetua se është e mundur të kryhet një tranzicion strukturor i disa polinukleotideve me dy fije homopurine-homopirimidine me formimin e një strukture me tre fije që përmban:
- një fije homopurine dhe dy homopirimidine ( Tripleksi Py-Pu-Py) [ndërveprim Hoogsteen].
Blloqet përbërëse të trefishit Py-Pu-Py janë treshe kanonike izomorfe CGC+ dhe TAT. Stabilizimi i tripleksit kërkon protonizimin e treshes CGC+, kështu që këto triplekse varen nga pH e tretësirës.
- një homopirimidinë dhe dy fije homopurine ( Tripleksi Py-Pu-Pu) [ndërveprim i anasjelltë i Hoogsteen-it].
Blloqet përbërëse të trefishit Py-Pu-Pu janë treshe kanonike izomorfe CGG dhe TAA. Një veti thelbësore e triplekseve Py-Pu-Pu është varësia e qëndrueshmërisë së tyre nga prania e joneve të dyfishtë të ngarkuar, dhe jone të ndryshëm nevojiten për të stabilizuar triplekset e sekuencave të ndryshme. Meqenëse formimi i treplekseve Py-Pu-Pu nuk kërkon protonimin e nukleotideve përbërëse të tyre, treplekse të tilla mund të ekzistojnë në pH neutral.
Shënim: ndërveprimet direkte dhe të kundërta Hoogsteen shpjegohen me simetrinë e 1-metiltiminës: një rrotullim prej 180° rezulton që atomi O2 të zërë vendin e atomit O4, ndërsa sistemi i lidhjeve hidrogjenore ruhet.
Njihen dy lloje të helikave të trefishta:
- spirale të trefishta paralele në të cilat polariteti i vargut të tretë përkon me polaritetin e zinxhirit homopurin të dupleksit Watson-Crick
- spirale të trefishta antiparalele, në të cilat polaritetet e zinxhirit të tretë dhe homopurinës janë të kundërta.

G-katërfish- ADN me 4 vargje. Kjo strukturë formohet nëse ka katër guanina, të cilat formojnë të ashtuquajturin G-quadruplex - një valle e rrumbullakët e katër guaninave.
Lëvizjet e para për mundësinë e formimit të strukturave të tilla u morën shumë përpara punës së përparimit të Watson dhe Crick - në vitin 1910. Pastaj kimisti gjerman Ivar Bang zbuloi se një nga përbërësit e ADN-së - acidi guanosinik - formon xhel në përqendrime të larta, ndërsa përbërësit e tjerë të ADN-së nuk e kanë këtë veti.
Në vitin 1962, duke përdorur metodën e difraksionit me rreze X, u bë e mundur të vendoset struktura qelizore e këtij xhel. Doli të ishte i përbërë nga katër mbetje guanine, që lidhnin njëra-tjetrën në një rreth dhe formonin një katror karakteristik. Në qendër, lidhja mbështetet nga një jon metalik (Na, K, Mg). Të njëjtat struktura mund të formohen në ADN nëse përmban shumë guaninë. Këta katrorë të rrafshët (Kuartetet G) grumbullohen për të formuar struktura mjaft të qëndrueshme dhe të dendura (G-katërplekse).
Katër fije të veçanta të ADN-së mund të thuren në komplekse me katër fije, por ky është më tepër një përjashtim. Më shpesh, një fije e vetme e acidit nukleik thjesht lidhet në një nyjë, duke formuar trashje karakteristike (për shembull, në skajet e kromozomeve), ose ADN me dy fije në disa rajone të pasura me guaninë formon një katërfish lokal.
Ekzistenca e katërfishtë në skajet e kromozomeve - në telomere dhe në nxitësit e tumorit - është studiuar më së shumti. Megjithatë, ende nuk dihet një pamje e plotë e lokalizimit të ADN-së së tillë në kromozomet e njeriut.
Të gjitha këto struktura të pazakonta të ADN-së në formë lineare janë të paqëndrueshme në krahasim me ADN-në e formës B. Sidoqoftë, ADN-ja shpesh ekziston në një formë rrethore të tensionit topologjik kur ka atë që quhet mbimbështjellje. Në këto kushte, strukturat jo-kanonike të ADN-së formohen lehtësisht: format Z, "kryqet" dhe "furkët", format H, katërfishat e guaninës dhe i-motivi.
- Forma e mbimbështjellur - vihet re kur lirohet nga bërthama e qelizës pa dëmtuar shtyllën kurrizore të pentozofosfatit. Ka formën e unazave të mbyllura super të përdredhura. Në gjendjen e mbimbështjellur, spiralen e dyfishtë e ADN-së "përdredhet në vetvete" të paktën një herë, domethënë përmban të paktën një mbikthim (merr formën e një figure tetë).
- Gjendja e relaksuar e ADN-së - vërehet me një thyerje të vetme (thyerja e një fije floku). Në këtë rast, superbobinat zhduken dhe ADN-ja merr formën e një unaze të mbyllur.
- Forma lineare e ADN-së vërehet kur dy fijet e një spirale të dyfishtë thyhen.
Struktura terciare e ADN-së
Struktura terciare e ADN-së formohet si rezultat i përdredhjes shtesë në hapësirën e një molekule me dy spirale - mbimbështjellja e saj. Mbimbështjellja e molekulës së ADN-së në qelizat eukariote, ndryshe nga prokariotët, ndodh në formën e komplekseve me proteina.
Pothuajse e gjithë ADN-ja e eukarioteve gjendet në kromozomet e bërthamave; vetëm një sasi e vogël gjendet në mitokondri dhe në bimë në plastide. Substanca kryesore e kromozomeve të qelizave eukariotike (përfshirë kromozomet njerëzore) është kromatina, e përbërë nga ADN me dy fije, proteina histonike dhe johistone.
Proteinat e kromatinës histonike
Histonet janë proteina të thjeshta që përbëjnë deri në 50% të kromatinës. Në të gjitha qelizat shtazore dhe bimore të studiuara, u gjetën pesë klasa kryesore të histoneve: H1, H2A, H2B, H3, H4, të ndryshme në madhësi, përbërje aminoacide dhe ngarkesë (gjithmonë pozitive).
Histoni H1 i gjitarëve përbëhet nga një zinxhir i vetëm polipeptid që përmban afërsisht 215 aminoacide; madhësitë e histoneve të tjera variojnë nga 100 në 135 aminoacide. Të gjithë ata janë të spiralizuar dhe të përdredhur në një rruzull me një diametër prej rreth 2.5 nm dhe përmbajnë një sasi jashtëzakonisht të madhe të aminoacideve të ngarkuar pozitivisht lizinën dhe argininën. Histonet mund të acetilohen, metilohen, fosforilohen, poli(ADP)-ribozilohen dhe histonet H2A dhe H2B janë të lidhura në mënyrë kovalente me ubiquitin. Roli i modifikimeve të tilla në formimin e strukturës dhe kryerjen e funksioneve nga histonet ende nuk është sqaruar plotësisht. Supozohet se kjo është aftësia e tyre për të bashkëvepruar me ADN-në dhe për të siguruar një nga mekanizmat për rregullimin e veprimit të gjeneve.
Histonet ndërveprojnë me ADN-në kryesisht përmes lidhjeve jonike (urave të kripës) të formuara midis grupeve të fosfatit të ADN-së të ngarkuar negativisht dhe mbetjeve të histoneve të lizinës dhe argininës të ngarkuara pozitivisht.
Proteinat e kromatinës jo-histone
Proteinat johistone, ndryshe nga histonet, janë shumë të ndryshme. Janë izoluar deri në 590 fraksione të ndryshme të proteinave johistone që lidhen me ADN-në. Ato quhen gjithashtu proteina acidike, pasi struktura e tyre dominohet nga aminoacide acidike (ato janë polianione). Diversiteti i proteinave jo-histone shoqërohet me rregullimin specifik të aktivitetit të kromatinës. Për shembull, enzimat e nevojshme për replikimin dhe shprehjen e ADN-së mund të lidhen me kromatinën në mënyrë të përkohshme. Proteinat e tjera, të themi, ato të përfshira në procese të ndryshme rregullatore, lidhen me ADN-në vetëm në inde specifike ose në faza të caktuara të diferencimit. Çdo proteinë është plotësuese e një sekuence specifike të nukleotideve të ADN-së (vendndodhja e ADN-së). Ky grup përfshin:
- familja e proteinave të gishtave të zinkut të zonës specifike. Çdo "gisht zinku" njeh një vend specifik të përbërë nga 5 çifte nukleotide.
- familja e proteinave specifike për vendin - homodimerët. Fragmenti i një proteine të tillë në kontakt me ADN-në ka një strukturë spirale-kthesë-helix.
- Proteinat e xhelit me lëvizshmëri të lartë (proteinat HMG) janë një grup proteinash strukturore dhe rregullatore që shoqërohen vazhdimisht me kromatinë. Ato kanë një peshë molekulare më të vogël se 30 kDa dhe karakterizohen nga një përmbajtje e lartë e aminoacideve të ngarkuara. Për shkak të peshës së tyre të ulët molekulare, proteinat HMG kanë lëvizshmëri të lartë gjatë elektroforezës së xhelit të poliakrilamidit.
- enzimat e riprodhimit, transkriptimit dhe riparimit.
Me pjesëmarrjen e proteinave strukturore, rregullatore dhe enzimave të përfshira në sintezën e ADN-së dhe ARN-së, filli nukleozom shndërrohet në një kompleks shumë të kondensuar të proteinave dhe ADN-së. Struktura që rezulton është 10,000 herë më e shkurtër se molekula origjinale e ADN-së.

Kromatinë
Kromatina është një kompleks proteinash me ADN bërthamore dhe substanca inorganike. Pjesa më e madhe e kromatinës është joaktive. Ai përmban ADN të ngjeshur dhe të kondensuar. Kjo është heterokromatina. Ekzistojnë kromatinë konstituive, gjenetikisht joaktive (ADN satelitore) e përbërë nga rajone jo të shprehura, dhe fakultative - joaktive në një numër brezash, por në rrethana të caktuara të aftë për t'u shprehur.
Kromatina aktive (eukromatina) është e pakondensuar, d.m.th. paketuar më pak fort. Në qeliza të ndryshme përmbajtja e tij varion nga 2 deri në 11%. Në qelizat e trurit është më e bollshme - 10-11%, në qelizat e mëlçisë - 3-4 dhe qelizat e veshkave - 2-3%. Vërehet transkriptimi aktiv i eukromatinës. Për më tepër, organizimi i tij strukturor lejon që i njëjti informacion gjenetik i ADN-së, i natyrshëm në një lloj të caktuar organizmi, të përdoret ndryshe në qeliza të specializuara.
Në një mikroskop elektronik, imazhi i kromatinës i ngjan rruazave: trashje sferike rreth 10 nm në madhësi, të ndara nga ura si fije. Këto trashje sferike quhen nukleozome. Nukleozoma është një njësi strukturore e kromatinës. Çdo nukleozom përmban një segment të ADN-së të mbështjellë me 146 bp, duke formuar 1,75 kthesa majtas për bërthamën nukleozomale. Bërthama nukleozomale është një oktamer histoni i përbërë nga histonet H2A, H2B, H3 dhe H4, dy molekula të secilit lloj (Fig. 9), i cili duket si një disk me diametër 11 nm dhe trashësi 5,7 nm. Histoni i pestë, H1, nuk është pjesë e bërthamës nukleozomale dhe nuk është i përfshirë në procesin e mbështjelljes së ADN-së në oktamerin e histonit. Ai kontakton ADN-në në vendet ku spiralen e dyfishtë hyn dhe del nga bërthama nukleozomale. Këto janë seksione të ADN-së intercore (lidhëse), gjatësia e të cilave varion në varësi të llojit të qelizës nga 40 në 50 çifte nukleotide. Si rezultat, gjatësia e fragmentit të ADN-së të përfshirë në nukleozome gjithashtu ndryshon (nga 186 në 196 çifte nukleotide).
Nukleozomet përmbajnë afërsisht 90% ADN, pjesa tjetër janë lidhëse. Besohet se nukleozomet janë fragmente të kromatinës "të heshtur" dhe lidhësi është aktiv. Megjithatë, nukleozomet mund të shpalosen dhe të bëhen lineare. Nukleozomet e shpalosura janë tashmë kromatinë aktive. Kjo tregon qartë varësinë e funksionit nga struktura. Mund të supozohet se sa më shumë kromatinë të përmbahet në nukleozome globulare, aq më pak aktive është. Natyrisht, në qeliza të ndryshme proporcioni i pabarabartë i kromatinës në pushim është i lidhur me numrin e nukleozomeve të tilla.
Në fotografitë mikroskopike elektronike, në varësi të kushteve të izolimit dhe shkallës së shtrirjes, kromatina mund të duket jo vetëm si një fije e gjatë me trashje - "rruaza" nukleozomesh, por edhe si një fibril (fibër) më i shkurtër dhe më i dendur me një diametër prej 30 nm, formimi i të cilit vërehet gjatë ndërveprimit të histonit H1 i lidhur me rajonin lidhës të ADN-së dhe histonit H3, gjë që çon në përdredhje shtesë të spirales prej gjashtë nukleozomesh në kthesë për të formuar një solenoid me diametër 30 nm. Në këtë rast, proteina e histonit mund të ndërhyjë në transkriptimin e një numri gjenesh dhe në këtë mënyrë të rregullojë aktivitetin e tyre.
Si rezultat i ndërveprimeve të ADN-së me histonet e përshkruara më sipër, një segment i një spirale të dyfishtë të ADN-së prej 186 çifte bazash me një diametër mesatar prej 2 nm dhe një gjatësi prej 57 nm, shndërrohet në një spirale me diametër 10 nm dhe gjatësi prej 5 nm. Kur kjo spirale ngjeshet më pas në një fibër me diametër 30 nm, shkalla e kondensimit rritet gjashtëfish.
Në fund të fundit, paketimi i një dupleksi të ADN-së me pesë histone rezulton në kondensim 50-fish të ADN-së. Megjithatë, edhe një shkallë kaq e lartë e kondensimit nuk mund të shpjegojë ngjeshjen pothuajse 50,000 - 100,000-fish të ADN-së në kromozomin e metafazës. Fatkeqësisht, detajet e paketimit të mëtejshëm të kromatinës deri në kromozomin e metafazës nuk dihen ende, kështu që ne mund të konsiderojmë vetëm tiparet e përgjithshme të këtij procesi.
Nivelet e ngjeshjes së ADN-së në kromozome
Çdo molekulë e ADN-së paketohet në një kromozom të veçantë. Qelizat diploide njerëzore përmbajnë 46 kromozome, të cilat ndodhen në bërthamën e qelizës. Gjatësia totale e ADN-së së të gjitha kromozomeve në një qelizë është 1,74 m, por diametri i bërthamës në të cilën janë paketuar kromozomet është miliona herë më i vogël. Një paketim i tillë kompakt i ADN-së në kromozome dhe kromozome në bërthamën e qelizës sigurohet nga një shumëllojshmëri e proteinave histonike dhe johistone që ndërveprojnë në një sekuencë të caktuar me ADN-në (shih më lart). Ngjeshja e ADN-së në kromozome bën të mundur reduktimin e dimensioneve të saj lineare me afërsisht 10,000 herë - afërsisht nga 5 cm në 5 mikron. Ka disa nivele të ngjeshjes (Fig. 10).


- Spiralja e dyfishtë e ADN-së është një molekulë e ngarkuar negativisht me një diametër prej 2 nm dhe një gjatësi prej disa cm.
- niveli nukleozom- kromatina duket në një mikroskop elektronik si një zinxhir "rruazash" - nukleozome - "në një fije". Nukleozomi është një njësi strukturore universale që gjendet si në eukromatinë ashtu edhe në heterokromatinë, në bërthamën ndërfazore dhe kromozomet metafazë.
Niveli nukleozomik i ngjeshjes sigurohet nga proteinat speciale - histonet. Tetë domene histonike të ngarkuara pozitivisht formojnë bërthamën e nukleozomes rreth së cilës është plagosur një molekulë e ADN-së e ngarkuar negativisht. Kjo jep një shkurtim prej 7 herë, ndërsa diametri rritet nga 2 në 11 nm.
- niveli i solenoidit
Niveli solenoid i organizimit të kromozomeve karakterizohet nga përdredhja e filamentit nukleozom dhe formimi i fibrileve më të trasha me diametër 20-35 nm - solenoidë ose superbid. Lartësia e solenoidit është 11 nm; ka rreth 6-10 nukleozome në rrotullim. Paketimi solenoid konsiderohet më i mundshëm sesa paketimi superbid, sipas të cilit një fibril kromatin me diametër 20-35 nm është një zinxhir granulash, ose superbidësh, secila prej të cilave përbëhet nga tetë nukleozome. Në nivelin e solenoidit, madhësia lineare e ADN-së zvogëlohet me 6-10 herë, diametri rritet në 30 nm.
- niveli i lakut
Niveli i lakut sigurohet nga proteinat lidhëse të ADN-së jo-histone specifike të vendit që njohin dhe lidhen me sekuenca specifike të ADN-së, duke formuar unaza prej afërsisht 30-300 kb. Laku siguron shprehjen e gjeneve, d.m.th. laku nuk është vetëm një formacion strukturor, por edhe funksional. Shkurtimi në këtë nivel ndodh 20-30 herë. Diametri rritet në 300 nm. Në preparatet citologjike mund të shihen struktura në formë lakore si "brushat e llambave" në ovocitet e amfibëve. Këto sythe duket se janë të mbimbështjellura dhe përfaqësojnë domene të ADN-së, ndoshta që korrespondojnë me njësitë e transkriptimit dhe replikimit të kromatinës. Proteinat specifike rregullojnë bazat e sytheve dhe, ndoshta, disa nga seksionet e tyre të brendshme. Organizimi i domenit të ngjashëm me lak promovon palosjen e kromatinës në kromozomet metafazë në struktura spirale të rendit më të lartë.
- niveli i domenit
Niveli i domenit të organizimit të kromozomeve nuk është studiuar mjaftueshëm. Në këtë nivel vërehet formimi i domeneve të lakut - strukturat e fijeve (fibrileve) 25-30 nm të trasha, të cilat përmbajnë 60% proteina, 35% ADN dhe 5% ARN, janë praktikisht të padukshme në të gjitha fazat e ciklit qelizor me me përjashtim të mitozës dhe shpërndahen disi rastësisht në të gjithë bërthamën qelizore. Në preparatet citologjike mund të shihen struktura në formë lakore si "brushat e llambave" në ovocitet e amfibëve.
Domenet e lakut janë bashkangjitur në bazën e tyre në matricën e proteinave intranukleare në të ashtuquajturat vende të bashkëngjitjes së integruar, shpesh të referuara si sekuenca MAR/SAR (MAR, nga rajoni i lidhur me matricën angleze; SAR, nga rajonet e bashkëngjitjes së skelës angleze) - Fragmentet e ADN-së disa qindra çifte bazash në gjatësi që karakterizohen nga një përmbajtje e lartë (>65%) e çifteve të nukleotideve A/T. Çdo domen duket se ka një origjinë të vetme replikimi dhe funksionon si një njësi superhelike autonome. Çdo domen loop përmban shumë njësi transkriptimi, funksionimi i të cilave ka të ngjarë të jetë i koordinuar - i gjithë domeni është ose në gjendje aktive ose joaktive.
Në nivelin e domenit, si rezultat i paketimit sekuencial të kromatinës, një rënie në dimensionet lineare të ADN-së ndodh përafërsisht 200 herë (700 nm).
- niveli kromozomik
Në nivelin kromozomik, kondensimi i kromozomit profazë në një kromozom metafazë ndodh me ngjeshjen e domeneve të lakut rreth kornizës boshtore të proteinave johistone. Kjo mbimbështjellje shoqërohet me fosforilim të të gjitha molekulave H1 në qelizë. Si rezultat, kromozomi i metafazës mund të përshkruhet si sythe solenoide të mbushura dendur, të mbështjellë në një spirale të ngushtë. Një kromozom tipik i njeriut mund të përmbajë deri në 2600 sythe. Trashësia e një strukture të tillë arrin 1400 nm (dy kromatide), dhe molekula e ADN-së shkurtohet për 104 herë, d.m.th. nga 5 cm e shtrirë ADN-në në 5 μm.
Funksionet e kromozomeve
Në ndërveprim me mekanizmat ekstrakromozomalë, kromozomet sigurojnë
- ruajtja e informacionit trashëgues
- duke përdorur këtë informacion për të krijuar dhe mbajtur organizimin celular
- rregullimi i leximit të informacionit trashëgues
- vetë-dyfishimi i materialit gjenetik
- transferimi i materialit gjenetik nga qeliza amë në qelizat bijë.
Ka të dhëna se kur aktivizohet një rajon i kromatinës, d.m.th. gjatë transkriptimit, së pari histoni H1 dhe më pas okteti i histonit hiqen në mënyrë të kthyeshme prej tij. Kjo shkakton dekondensimin e kromatinës, kalimin sekuencial të një fibrili kromatin 30 nm në një fibril 10 nm dhe shpalosjen e mëtejshme të tij në seksione të ADN-së së lirë, d.m.th. humbja e strukturës së nukleozomeve.
Pas zbulimit të parimit të organizimit molekular të një substance të tillë si ADN-ja në 1953, filloi të zhvillohet biologjia molekulare. Më tej në procesin e kërkimit, shkencëtarët zbuluan se si rikombinohet ADN-ja, përbërja e saj dhe si është strukturuar gjenomi ynë njerëzor.
Çdo ditë ndodhin procese komplekse në nivel molekular. Si është e strukturuar molekula e ADN-së, nga çfarë përbëhet? Dhe çfarë roli luajnë molekulat e ADN-së në një qelizë? Le të flasim në detaje për të gjitha proceset që ndodhin brenda zinxhirit të dyfishtë.
Çfarë është informacioni trashëgues?
Pra, ku filloi gjithçka? Në vitin 1868 ata e gjetën atë në bërthamat e baktereve. Dhe në vitin 1928, N. Koltsov parashtroi teorinë se i gjithë informacioni gjenetik për një organizëm të gjallë është i koduar në ADN. Pastaj J. Watson dhe F. Crick gjetën një model për spiralen tashmë të njohur të ADN-së në vitin 1953, për të cilën me meritë morën njohjen dhe një çmim - Çmimin Nobel.
Çfarë është ADN gjithsesi? Kjo substancë përbëhet nga 2 fije të bashkuara, ose më mirë spirale. Një pjesë e një zinxhiri të tillë me informacion të caktuar quhet gjen.

ADN-ja ruan të gjithë informacionin se çfarë lloj proteinash do të formohen dhe në çfarë rendi. Makromolekula e ADN-së është një bartës material i informacionit tepër voluminoz, i cili regjistrohet në një sekuencë strikte të tullave individuale - nukleotide. Gjithsej janë 4 nukleotide; ato plotësojnë njëra-tjetrën kimikisht dhe gjeometrikisht. Ky parim i plotësimit, ose komplementaritetit, në shkencë do të përshkruhet më vonë. Ky rregull luan një rol kyç në kodimin dhe dekodimin e informacionit gjenetik.
Meqenëse vargu i ADN-së është tepër i gjatë, nuk ka përsëritje në këtë sekuencë. Çdo krijesë e gjallë ka vargun e vet unik të ADN-së.
Funksionet e ADN-së
Funksionet përfshijnë ruajtjen e informacionit të trashëguar dhe transmetimin e tij tek pasardhësit. Pa këtë funksion, gjenomi i një specie nuk mund të ruhej dhe të zhvillohej gjatë mijëra viteve. Organizmat që kanë pësuar mutacione të rënda të gjeneve kanë më shumë gjasa të mos mbijetojnë ose të humbasin aftësinë për të prodhuar pasardhës. Kështu ndodh mbrojtja natyrore kundër degjenerimit të specieve.

Një funksion tjetër thelbësor është zbatimi i informacionit të ruajtur. Një qelizë nuk mund të krijojë një proteinë të vetme vitale pa ato udhëzime që ruhen në një zinxhir të dyfishtë.
Përbërja e acidit nukleik
Tani dihet me siguri se nga përbëhen vetë nukleotidet - blloqet ndërtuese të ADN-së. Ato përmbajnë 3 substanca:
- Acidi ortofosforik.
- Baza azotike. Bazat pirimidine - të cilat kanë vetëm një unazë. Këto përfshijnë timinë dhe citozinë. Bazat purine, të cilat përmbajnë 2 unaza. Këto janë guanina dhe adenina.
- Saharoza. ADN-ja përmban deoksiribozë, ARN-ja përmban ribozë.
Numri i nukleotideve është gjithmonë i barabartë me numrin e bazave azotike. Në laboratorë të veçantë, nukleotidi zbërthehet dhe baza azotike izolohet prej tij. Kështu studiohen vetitë individuale të këtyre nukleotideve dhe mutacionet e mundshme në to.
Nivelet e organizimit të informacionit trashëgues
Ekzistojnë 3 nivele organizimi: gjenetik, kromozomik dhe gjenomik. I gjithë informacioni i nevojshëm për sintezën e një proteine të re gjendet në një pjesë të vogël të zinxhirit - gjeni. Kjo do të thotë, gjeni konsiderohet si niveli më i ulët dhe më i thjeshtë i kodimit të informacionit.

Gjenet, nga ana tjetër, grumbullohen në kromozome. Falë këtij organizimi të bartësit të materialit trashëgues, grupet e karakteristikave alternojnë sipas ligjeve të caktuara dhe transmetohen nga një brez në tjetrin. Duhet theksuar se në trup ka një numër të pabesueshëm gjenesh, por informacioni nuk humbet edhe kur rikombinohet shumë herë.
Ka disa lloje gjenesh:
- Sipas qëllimit të tyre funksional, ekzistojnë 2 lloje: sekuenca strukturore dhe rregullatore;
- Bazuar në ndikimin e tyre në proceset që ndodhin në qelizë, ata dallojnë: gjenet supervitale, vdekjeprurëse, me kusht vdekjeprurëse, si dhe gjenet mutator dhe antimutator.
Gjenet janë të rregulluar përgjatë kromozomit në një rend linear. Në kromozome, informacioni nuk përqendrohet rastësisht; ekziston një renditje e caktuar. Madje ekziston edhe një hartë që tregon pozicionet ose vendndodhjet e gjeneve. Për shembull, dihet se kromozomi nr. 18 kodon të dhënat për ngjyrën e syve të një fëmije.
Çfarë është një gjenom? Ky është emri i dhënë për të gjithë grupin e sekuencave nukleotide në qelizën e një organizmi. Gjenomi karakterizon një specie të tërë, jo një individ.
Cili është kodi gjenetik i njeriut?
Fakti është se i gjithë potenciali i madh i zhvillimit njerëzor qëndron tashmë në periudhën e konceptimit. Të gjitha informacionet trashëgimore që janë të nevojshme për zhvillimin e zigotit dhe rritjen e fëmijës pas lindjes janë të koduara në gjene. Seksionet e ADN-së janë bartësit më bazë të informacionit trashëgues.
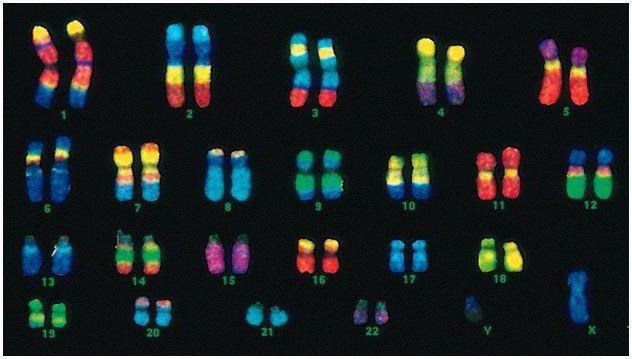
Njerëzit kanë 46 kromozome, ose 22 çifte somatike plus një kromozom që përcakton seksin nga secili prind. Ky grup diploid kromozomesh kodon të gjithë pamjen fizike të një personi, aftësitë e tij mendore dhe fizike dhe ndjeshmërinë ndaj sëmundjeve. Kromozomet somatike janë të padallueshme nga jashtë, por ato përmbajnë informacione të ndryshme, pasi njëri prej tyre është nga babai, tjetri nga nëna.
Kodi mashkullor ndryshon nga kodi femëror në çiftin e fundit të kromozomeve - XY. Seti diploid femëror është çifti i fundit, XX. Meshkujt marrin një kromozom X nga nëna e tyre biologjike, i cili më pas u kalohet vajzave të tyre. Kromozomi i seksit Y u kalon djemve.
Kromozomet e njeriut ndryshojnë shumë në madhësi. Për shembull, çifti më i vogël i kromozomeve është numri 17. Dhe çifti më i madh është 1 dhe 3.
Diametri i spirales së dyfishtë tek njerëzit është vetëm 2 nm. ADN-ja është e mbështjellë aq fort saqë përshtatet brenda bërthamës së vogël të një qelize, megjithëse do të ishte deri në 2 metra e gjatë nëse nuk përdredhohej. Gjatësia e spirales është qindra miliona nukleotide.
Si transmetohet kodi gjenetik?
Pra, çfarë roli luajnë molekulat e ADN-së në ndarjen e qelizave? Gjenet - bartës të informacionit trashëgues - ndodhen brenda çdo qelize të trupit. Për të kaluar kodin e tyre te një organizëm bijë, shumë krijesa ndajnë ADN-në e tyre në 2 spirale identike. Kjo quhet përsëritje. Gjatë procesit të replikimit, ADN-ja zbërthehet dhe "makinat" speciale plotësojnë secilën fije. Pasi heliksi gjenetik bifurkohet, bërthama dhe të gjitha organelet fillojnë të ndahen, dhe më pas e gjithë qeliza.
Por njerëzit kanë një proces tjetër të transmetimit të gjeneve - seksual. Karakteristikat e babait dhe nënës janë të përziera, kodi i ri gjenetik përmban informacione nga të dy prindërit.
Ruajtja dhe transmetimi i informacionit trashëgues është i mundur për shkak të organizimit kompleks të spirales së ADN-së. Në fund të fundit, siç thamë, struktura e proteinave është e koduar në gjene. Pasi të krijohet në kohën e konceptimit, ky kod do të kopjohet gjatë gjithë jetës. Kariotipi (bashkësia personale e kromozomeve) nuk ndryshon gjatë rinovimit të qelizave të organeve. Transferimi i informacionit kryhet me ndihmën e gameteve seksuale - mashkull dhe femër.
Vetëm viruset që përmbajnë një varg të ARN-së nuk janë në gjendje të transmetojnë informacionin e tyre tek pasardhësit e tyre. Prandaj, ata kanë nevojë për qeliza njerëzore ose shtazore për t'u riprodhuar.
Zbatimi i informacionit trashëgues
Në bërthamën e qelizës ndodhin vazhdimisht procese të rëndësishme. I gjithë informacioni i regjistruar në kromozome përdoret për të ndërtuar proteina nga aminoacidet. Por zinxhiri i ADN-së nuk largohet kurrë nga bërthama, kështu që ka nevojë për ndihmën e një përbërësi tjetër të rëndësishëm: ARN-së. Është ARN që është në gjendje të depërtojë në membranën bërthamore dhe të ndërveprojë me zinxhirin e ADN-së.
Nëpërmjet bashkëveprimit të ADN-së dhe 3 llojeve të ARN-së realizohet i gjithë informacioni i koduar. Në çfarë niveli ndodh zbatimi i informacionit trashëgues? Të gjitha ndërveprimet ndodhin në nivelin e nukleotideve. ARN-ja e dërguar kopjon një pjesë të zinxhirit të ADN-së dhe e sjell këtë kopje në ribozom. Këtu fillon sinteza e një molekule të re nga nukleotidet.
Në mënyrë që mRNA të kopjojë pjesën e nevojshme të zinxhirit, spiralen shpaloset dhe më pas, pas përfundimit të procesit të rikodimit, restaurohet përsëri. Për më tepër, ky proces mund të ndodhë njëkohësisht në 2 anët e 1 kromozomi.
Parimi i komplementaritetit
Ato përbëhen nga 4 nukleotide - adenina (A), guanina (G), citozina (C), timina (T). Ato lidhen me lidhje hidrogjenore sipas rregullit të komplementaritetit. Puna e E. Chargaff ndihmoi në vendosjen e këtij rregulli, pasi shkencëtari vuri re disa modele në sjelljen e këtyre substancave. E. Chargaff zbuloi se raporti molar i adeninës ndaj timinës është i barabartë me një. Dhe në të njëjtën mënyrë, raporti i guaninës ndaj citozinës është gjithmonë i barabartë me një.
Bazuar në punën e tij, gjenetistët formuan një rregull për bashkëveprimin e nukleotideve. Rregulli i komplementaritetit thotë se adenina kombinohet vetëm me timinën, dhe guanina kombinohet vetëm me citozinën. Gjatë dekodimit të spirales dhe sintezës së një proteine të re në ribozom, ky rregull i alternimit ndihmon për të gjetur shpejt aminoacidin e nevojshëm që është ngjitur në ARN-në e transferimit.
ARN dhe llojet e saj
Çfarë është informacioni trashëgues? nukleotidet në një varg të dyfishtë të ADN-së. Çfarë është ARN? Cila është puna e saj? ARN, ose acidi ribonukleik, ndihmon në nxjerrjen e informacionit nga ADN-ja, dekodimin e tij dhe, bazuar në parimin e komplementaritetit, në krijimin e proteinave të nevojshme për qelizat.
Gjithsej ekzistojnë 3 lloje të ARN-së. Secila prej tyre kryen rreptësisht funksionin e vet.
- Informacione (mRNA), ose quhet edhe matricë. Ai shkon drejt e në qendër të qelizës, në bërthamë. Gjen në një nga kromozomet materialin gjenetik të nevojshëm për të ndërtuar një proteinë dhe kopjon njërën nga anët e fijes së dyfishtë. Kopjimi ndodh sërish sipas parimit të komplementaritetit.
- Transportiështë një molekulë e vogël që ka dekodera nukleotide në njërën anë, dhe aminoacide që korrespondojnë me kodin bazë në anën tjetër. Detyra e tRNA është ta dërgojë atë në "punëtori", domethënë në ribozom, ku sintetizon aminoacidin e nevojshëm.
- rRNA është ribozomale. Ai kontrollon sasinë e proteinave që prodhohet. Ai përbëhet nga 2 pjesë - një seksion aminoacid dhe një seksion peptid.
I vetmi ndryshim në dekodim është se ARN nuk ka timinë. Në vend të timinës, uracili është i pranishëm këtu. Por më pas, gjatë procesit të sintezës së proteinave, tRNA ende instalon saktë të gjitha aminoacidet. Nëse ndodh ndonjë dështim në dekodimin e informacionit, atëherë ndodh një mutacion.
Riparimi i molekulës së dëmtuar të ADN-së
Procesi i rivendosjes së një fije të dyfishtë të dëmtuar quhet riparim. Gjatë procesit të riparimit, gjenet e dëmtuara hiqen.

Pastaj sekuenca e kërkuar e elementeve riprodhohet saktësisht dhe pritet përsëri në të njëjtin vend në zinxhirin nga ku është hequr. E gjithë kjo ndodh falë kimikateve të veçanta - enzimave.
Pse ndodhin mutacionet?
Pse disa gjene fillojnë të ndryshojnë dhe pushojnë së kryeri funksionin e tyre - duke ruajtur informacionin jetik të trashëguar? Kjo ndodh për shkak të një gabimi në dekodim. Për shembull, nëse adenina zëvendësohet aksidentalisht me timinë.
Ekzistojnë gjithashtu mutacione kromozomike dhe gjenomike. Mutacionet kromozomale ndodhin kur seksionet e informacionit të trashëguar humbasin, dyfishohen ose madje transferohen dhe futen në një kromozom tjetër.

Mutacionet gjenomike janë më seriozet. Shkaku i tyre është një ndryshim në numrin e kromozomeve. Kjo do të thotë, kur në vend të një çifti - një grup diploid, një grup triploid është i pranishëm në kariotip.
Shembulli më i famshëm i një mutacioni triploid është sindroma Down, në të cilën grupi personal i kromozomeve është 47. Në fëmijë të tillë, në vend të çiftit të 21-të formohen 3 kromozome.
Ekziston edhe një mutacion i njohur i quajtur poliploidi. Por poliploidia ndodh vetëm te bimët.
Molekula e ADN-së përbëhet nga dy vargje që formojnë një spirale të dyfishtë. Struktura e saj u deshifrua për herë të parë nga Francis Crick dhe James Watson në 1953.
Në fillim, molekula e ADN-së, e përbërë nga një palë zinxhirësh nukleotide të përdredhura rreth njëri-tjetrit, krijoi pyetje se pse kishte këtë formë të veçantë. Shkencëtarët e quajnë këtë fenomen komplementaritet, që do të thotë se vetëm disa nukleotide mund të gjenden përballë njëri-tjetrit në fijet e tij. Për shembull, adenina është gjithmonë përballë timinës, dhe guanina është gjithmonë e kundërta e citozinës. Këto nukleotide të molekulës së ADN-së quhen komplementare.
Skematikisht përshkruhet kështu:
T - A
C - G
Këto çifte formojnë një lidhje kimike nukleotide, e cila përcakton rendin e aminoacideve. Në rastin e parë është pak më e dobët. Lidhja midis C dhe G është më e fortë. Nukleotidet jo komplementare nuk formojnë çifte me njëri-tjetrin.
Rreth ndërtesës
Pra, struktura e molekulës së ADN-së është e veçantë. Ajo ka këtë formë për një arsye: fakti është se numri i nukleotideve është shumë i madh dhe nevojitet shumë hapësirë për të akomoduar zinxhirë të gjatë. Është për këtë arsye që zinxhirët karakterizohen nga një kthesë spirale. Ky fenomen quhet spiralizimi, ai lejon që fijet të shkurtohen me rreth pesë deri në gjashtë herë.
Trupi përdor disa molekula të këtij lloji në mënyrë shumë aktive, të tjera rrallë. Këta të fundit, përveç spiralizimit, i nënshtrohen edhe "paketimit kompakt" si superspiralizimi. Dhe pastaj gjatësia e molekulës së ADN-së zvogëlohet me 25-30 herë.

Çfarë është "paketimi" i një molekule?
Procesi i mbimbështjelljes përfshin proteinat e histonit. Ata kanë strukturën dhe pamjen e një bobine filli ose një shufre. Mbi to janë mbështjellë fijet e spiralizuara, të cilat menjëherë bëhen "të paketuara në mënyrë kompakte" dhe zënë pak hapësirë. Kur lind nevoja për të përdorur një ose një fije tjetër, ajo zgjidhet nga një bobinë, për shembull, një proteinë histoni, dhe spiralen hapet në dy zinxhirë paralelë. Kur molekula e ADN-së është në këtë gjendje, të dhënat e nevojshme gjenetike mund të lexohen prej saj. Megjithatë, ekziston një kusht. Marrja e informacionit është e mundur vetëm nëse struktura e molekulës së ADN-së ka një formë jo të përdredhur. Kromozomet që janë të arritshme për lexim quhen eukromatina, dhe nëse janë të mbimbështjellura, atëherë ato tashmë janë heterokromatina.
Acidet nukleike
Acidet nukleike, si proteinat, janë biopolimere. Funksioni kryesor është ruajtja, zbatimi dhe transmetimi i informacionit trashëgues (gjenetik). Ato vijnë në dy lloje: ADN dhe ARN (deoksiribonukleike dhe ribonukleike). Monomerët në to janë nukleotide, secila prej të cilave përmban një mbetje të acidit fosforik, një sheqer me pesë karbon (deoksiribozë/ribozë) dhe një bazë azotike. Kodi i ADN-së përfshin 4 lloje të nukleotideve - adeninë (A) / guaninë (G) / citozinë (C) / timinë (T). Ato ndryshojnë në bazën azotike që përmbajnë.
Në një molekulë të ADN-së, numri i nukleotideve mund të jetë i madh - nga disa mijëra në dhjetëra e qindra miliona. Molekula të tilla gjigante mund të ekzaminohen përmes një mikroskopi elektronik. Në këtë rast, do të mund të shihni një zinxhir të dyfishtë të fijeve polinukleotide, të cilat janë të lidhura me njëra-tjetrën me lidhje hidrogjenore të bazave azotike të nukleotideve.

Hulumtimi
Gjatë rrjedhës së hulumtimit, shkencëtarët zbuluan se llojet e molekulave të ADN-së ndryshojnë në organizma të ndryshëm të gjallë. U zbulua gjithashtu se guanina e një zinxhiri mund të lidhet vetëm me citozinën, dhe timina me adeninën. Rregullimi i nukleotideve në një zinxhir korrespondon rreptësisht me atë paralel. Falë këtij komplementariteti të polinukleotideve, molekula e ADN-së është e aftë të dyfishohet dhe të riprodhohet vetë. Por së pari, zinxhirët plotësues, nën ndikimin e enzimave të veçanta që shkatërrojnë nukleotidet e çiftuara, ndryshojnë dhe më pas në secilën prej tyre fillon sinteza e zinxhirit që mungon. Kjo ndodh për shkak të nukleotideve të lira të pranishme në sasi të mëdha në çdo qelizë. Si rezultat i kësaj, në vend të "molekulës së nënës", formohen dy "bijë", identike në përbërje dhe strukturë, dhe kodi i ADN-së bëhet ai origjinal. Ky proces është një pararendës i ndarjes së qelizave. Siguron transmetimin e të gjitha të dhënave trashëgimore nga qelizat e nënës tek qelizat bija, si dhe tek të gjitha gjeneratat pasuese.
Si lexohet kodi i gjeneve?
Sot, llogaritet jo vetëm masa e një molekule të ADN-së - është gjithashtu e mundur të zbulohen të dhëna më komplekse që më parë ishin të paarritshme për shkencëtarët. Për shembull, mund të lexoni informacione se si një organizëm përdor qelizën e tij. Natyrisht, në fillim ky informacion është në formë të koduar dhe ka formën e një matrice të caktuar, dhe për këtë arsye duhet të transportohet në një bartës të veçantë, që është ARN. Acidi ribonukleik është në gjendje të depërtojë në qelizë përmes membranës bërthamore dhe të lexojë informacionin e koduar brenda. Kështu, ARN është një bartës i të dhënave të fshehura nga bërthama në qelizë, dhe ndryshon nga ADN-ja në atë që përmban ribozë në vend të deoksiribozës dhe uracil në vend të timinës. Për më tepër, ARN është me një fije floku.

Sinteza e ARN-së
Analiza e thelluar e ADN-së ka treguar se pasi ARN del nga bërthama, ajo hyn në citoplazmë, ku mund të integrohet si matricë në ribozome (sisteme enzimë speciale). Të udhëhequr nga informacioni i marrë, ata mund të sintetizojnë sekuencën e duhur të aminoacideve proteinike. Ribozomi mëson nga kodi i trefishtë se cili lloj i përbërjes organike duhet të lidhet me zinxhirin e proteinave formuese. Çdo aminoacid ka trefishin e tij specifik, i cili e kodon atë.
Pas përfundimit të formimit të zinxhirit, ai merr një formë specifike hapësinore dhe shndërrohet në një proteinë të aftë për të kryer funksionet e saj hormonale, ndërtimore, enzimatike dhe të tjera. Për çdo organizëm është një produkt gjen. Është prej saj që përcaktohen të gjitha llojet e cilësive, vetive dhe manifestimeve të gjeneve.

Gjenet
Proceset e sekuencës u zhvilluan kryesisht për të marrë informacion se sa gjene ka një molekulë e ADN-së në strukturën e saj. Dhe, megjithëse kërkimet i kanë lejuar shkencëtarët të bëjnë përparim të madh në këtë çështje, ende nuk është e mundur të dihet numri i tyre i saktë.
Vetëm pak vite më parë supozohej se molekulat e ADN-së përmbajnë afërsisht 100 mijë gjene. Pak më vonë, shifra u ul në 80 mijë, dhe në vitin 1998, gjenetistët deklaruan se vetëm 50 mijë gjene janë të pranishëm në një ADN, të cilat janë vetëm 3% e gjatësisë totale të ADN-së. Por përfundimet e fundit të gjenetistëve ishin befasuese. Tani ata pretendojnë se gjenomi përfshin 25-40 mijë nga këto njësi. Rezulton se vetëm 1.5% e ADN-së kromozomale është përgjegjëse për kodimin e proteinave.
Hulumtimi nuk u ndal me kaq. Një ekip paralel specialistësh të inxhinierisë gjenetike zbuloi se numri i gjeneve në një molekulë është saktësisht 32 mijë. Siç mund ta shihni, është ende e pamundur të merret një përgjigje përfundimtare. Ka shumë kontradikta. Të gjithë studiuesit mbështeten vetëm në rezultatet e tyre.
A kishte evolucion?
Përkundër faktit se nuk ka prova të evolucionit të molekulës (pasi struktura e molekulës së ADN-së është e brishtë dhe e vogël në madhësi), shkencëtarët ende bënë një supozim. Bazuar në të dhënat laboratorike, ata shprehën versionin e mëposhtëm: në fazën fillestare të shfaqjes së saj, molekula kishte formën e një peptidi të thjeshtë vetë-përsëritës, i cili përfshinte deri në 32 aminoacide të gjetura në oqeanet e lashta.
Pas vetë-përsëritjes, në sajë të forcave të përzgjedhjes natyrore, molekulat fituan aftësinë për të mbrojtur veten nga elementët e jashtëm. Ata filluan të jetojnë më gjatë dhe të riprodhohen në sasi më të mëdha. Molekulat që u gjendën në flluskën e lipideve kishin çdo shans për të riprodhuar veten. Si rezultat i një sërë ciklesh të njëpasnjëshme, flluskat e lipideve fituan formën e membranave qelizore, dhe më pas - grimcat e njohura. Duhet të theksohet se sot çdo seksion i një molekule të ADN-së është një strukturë komplekse dhe qartësisht funksionale, të gjitha tiparet e së cilës shkencëtarët nuk i kanë studiuar ende plotësisht.

Bota moderne
Kohët e fundit, shkencëtarët nga Izraeli kanë zhvilluar një kompjuter që mund të kryejë triliona operacione në sekondë. Sot është makina më e shpejtë në Tokë. I gjithë sekreti është se pajisja inovative mundësohet nga ADN-ja. Profesorët thonë se në të ardhmen e afërt, kompjuterë të tillë do të mund të gjenerojnë edhe energji.
Një vit më parë, specialistë nga Instituti Weizmann në Rehovot (Izrael) njoftuan krijimin e një makinerie llogaritëse molekulare të programueshme të përbërë nga molekula dhe enzima. Ata zëvendësuan mikroçipet e silikonit me to. Deri më sot, skuadra ka bërë progres të mëtejshëm. Tani vetëm një molekulë e ADN-së mund t'i sigurojë një kompjuteri të dhënat e nevojshme dhe karburantin e nevojshëm.
"Nanokompjuterët" biokimikë nuk janë një trillim; ata tashmë ekzistojnë në natyrë dhe manifestohen në çdo krijesë të gjallë. Por shpesh ato nuk menaxhohen nga njerëzit. Një person nuk mund të operojë ende në gjenomin e ndonjë bime për të llogaritur, të themi, numrin "Pi".
Ideja e përdorimit të ADN-së për ruajtjen/përpunimin e të dhënave erdhi për herë të parë në mendjet e shkencëtarëve në 1994. Ishte atëherë që një molekulë u përdor për të zgjidhur një problem të thjeshtë matematikor. Që atëherë, një numër grupesh kërkimore kanë propozuar projekte të ndryshme në lidhje me kompjuterët e ADN-së. Por këtu të gjitha përpjekjet bazoheshin vetëm në molekulën e energjisë. Ju nuk mund ta shihni një kompjuter të tillë me sy të lirë; duket si një zgjidhje transparente uji në një provëz. Nuk ka pjesë mekanike në të, por vetëm triliona pajisje biomolekulare - dhe kjo është vetëm në një pikë lëngu!
ADN-ja e njeriut
Njerëzit u ndërgjegjësuan për llojin e ADN-së njerëzore në vitin 1953, kur shkencëtarët ishin në gjendje për herë të parë t'i demonstronin botës një model të ADN-së me dy fije. Për këtë, Kirk dhe Watson morën çmimin Nobel, pasi ky zbulim u bë themelor në shekullin e 20-të.
Me kalimin e kohës, natyrisht, ata vërtetuan se një molekulë e strukturuar njerëzore mund të duket jo vetëm si në versionin e propozuar. Pas kryerjes së një analize më të detajuar të ADN-së, ata zbuluan formën A-, B- dhe të majtën Z-. Forma A- është shpesh një përjashtim, pasi formohet vetëm nëse ka mungesë lagështie. Por kjo është e mundur vetëm në studimet laboratorike; për mjedisin natyror kjo është anormale; një proces i tillë nuk mund të ndodhë në një qelizë të gjallë.
Forma B është klasike dhe njihet si një zinxhir i dyfishtë me dorën e djathtë, por forma Z jo vetëm që është e përdredhur në drejtim të kundërt me të majtën, por gjithashtu ka një pamje më zigzag. Shkencëtarët kanë identifikuar edhe formën G-quadruplex. Struktura e saj nuk ka 2, por 4 fije. Sipas gjenetistëve, kjo formë shfaqet në zonat ku ka një sasi të tepërt të guaninës.

ADN artificiale
Sot tashmë ekziston ADN artificiale, e cila është një kopje identike e asaj reale; ajo ndjek në mënyrë të përkryer strukturën e spirales së dyfishtë natyrore. Por, ndryshe nga polinukleotidi origjinal, ai artificial ka vetëm dy nukleotide shtesë.
Meqenëse dublimi u krijua në bazë të informacionit të marrë nga studime të ndryshme të ADN-së reale, ai gjithashtu mund të kopjohet, të vetë-përsëritet dhe të zhvillohet. Ekspertët kanë punuar për krijimin e një molekule të tillë artificiale për rreth 20 vjet. Rezultati është një shpikje e mahnitshme që mund të përdorë kodin gjenetik në të njëjtën mënyrë si ADN-ja natyrore.
Katër bazave ekzistuese azotike, gjenetistët shtuan dy të tjera, të cilat u krijuan nga modifikimi kimik i bazave natyrore. Ndryshe nga ADN-ja natyrore, ADN-ja artificiale doli të ishte mjaft e shkurtër. Ai përmban vetëm 81 çifte bazash. Megjithatë, ai gjithashtu riprodhohet dhe evoluon.
Replikimi i një molekule të përftuar artificialisht bëhet falë reaksionit zinxhir të polimerazës, por deri më tani kjo nuk ndodh në mënyrë të pavarur, por me ndërhyrjen e shkencëtarëve. Ata në mënyrë të pavarur shtojnë enzimat e nevojshme në ADN-në e përmendur, duke e vendosur atë në një medium të lëngshëm të përgatitur posaçërisht.
Rezultati përfundimtar
Procesi dhe rezultati përfundimtar i zhvillimit të ADN-së mund të ndikohet nga faktorë të ndryshëm, siç janë mutacionet. Kjo e bën të nevojshme studimin e mostrave të materies në mënyrë që rezultati i analizës të jetë i besueshëm dhe i besueshëm. Një shembull është testi i atësisë. Por ne nuk mund të mos gëzohemi që incidente të tilla si mutacioni janë të rralla. Megjithatë, mostrat e materies rikontrollohen gjithmonë për të marrë informacion më të saktë bazuar në analizat.

ADN-ja e bimëve
Falë teknologjive me sekuencë të lartë (HTS), është bërë një revolucion në fushën e gjenomikës - është e mundur edhe nxjerrja e ADN-së nga bimët. Natyrisht, marrja e ADN-së me peshë molekulare me cilësi të lartë nga materiali bimor paraqet disa vështirësi për shkak të numrit të madh të kopjeve të ADN-së së mitokondrive dhe kloroplastit, si dhe nivelit të lartë të polisaharideve dhe përbërjeve fenolike. Për të izoluar strukturën që po shqyrtojmë në këtë rast, përdoren metoda të ndryshme.
Lidhja hidrogjenore në ADN
Lidhja e hidrogjenit në molekulën e ADN-së është përgjegjëse për tërheqjen elektromagnetike të krijuar midis një atomi hidrogjeni të ngarkuar pozitivisht që është i lidhur me një atom elektronegativ. Ky ndërveprim dipoli nuk plotëson kriterin e një lidhjeje kimike. Por mund të ndodhë në mënyrë ndërmolekulare ose në pjesë të ndryshme të molekulës, pra në mënyrë intramolekulare.

Një atom hidrogjeni i bashkëngjitet atomit elektronegativ që është dhuruesi i lidhjes. Një atom elektronegativ mund të jetë azoti, fluori ose oksigjeni. Ai - përmes decentralizimit - tërheq renë elektronike nga bërthama e hidrogjenit në vetvete dhe e bën atomin e hidrogjenit (pjesërisht) të ngarkuar pozitivisht. Meqenëse madhësia e H është e vogël në krahasim me molekulat dhe atomet e tjera, ngarkesa është gjithashtu e vogël.
Dekodimi i ADN-së
Para se të deshifrojnë një molekulë të ADN-së, shkencëtarët fillimisht marrin një numër të madh qelizash. Për punën më të saktë dhe më të suksesshme duhen rreth një milion prej tyre. Rezultatet e marra gjatë studimit krahasohen dhe regjistrohen vazhdimisht. Sot, dekodimi i gjenomit nuk është më një gjë e rrallë, por një procedurë e arritshme.

Sigurisht, deshifrimi i gjenomit të një qelize të vetme është një ushtrim jopraktik. Të dhënat e marra gjatë studimeve të tilla nuk janë me interes për shkencëtarët. Por është e rëndësishme të kuptohet se të gjitha metodat ekzistuese të deshifrimit, pavarësisht kompleksitetit të tyre, nuk janë mjaft efektive. Ata do të lejojnë vetëm leximin e 40-70% të ADN-së.
Megjithatë, profesorët e Harvardit njoftuan së fundmi një metodë përmes së cilës mund të deshifrohet 90% e gjenomit. Teknika bazohet në shtimin e molekulave të primerit në qelizat e izoluara, me ndihmën e të cilave fillon riprodhimi i ADN-së. Por edhe kjo metodë nuk mund të konsiderohet e suksesshme; ajo ende duhet të përpunohet përpara se të përdoret hapur në shkencë.
