DNK molekulasi joylashgan. DNK va genlar. DNKga yangi qarash
Irsiyatning biokimyoviy asoslari.
Nuklein kislotalarning genetik roli.
Nuklein kislotalar - ibtidoiydan murakkabgacha bo'lgan barcha hujayralarda joylashgan biologik polimerlar. Ular birinchi marta 1868 yilda Iogan Fridrix Misher tomonidan yadroviy materialga boy hujayralarda (leykotsitlar, qizil ikra sperma) kashf etilgan. "Nuklein kislotalar" atamasi 1889 yilda taklif qilingan.
Nuklein kislotalarning ikki turi mavjud: DNK, RNK (ATP - mononukleotid). DNK va RNK molekulalar - matritsalardir. DNK taxminan 6 * 10 -12 g somatik hujayralarda mavjud: yadroda, mitoxondriyada. RNK ribosomalarning bir qismi bo'lib, yadro va sitoplazmada joylashgan.
Nuklein kislotalarning irsiy ma'lumotni uzatishdagi etakchi rolini o'rganish va isbotlash virusli zarralar ustida olib borildi. Tamaki mozaikasi virusi tamaki va chinor uchun xavfli ekanligi ma'lum. Virusli zarracha 95% oqsil va 5% nuklein kislotadan iborat. Virusli zarrachalardagi oqsil kapsidi almashtirildi, biroq bir muncha vaqt o'tgach, ikkala shtammdagi oqsil avvalgi shakliga aylandi.
E. coli ni yuqtirgan bakteriofaglarda fag qoplami oqsillari radioaktiv S bilan, fag DNKsi esa radioaktiv P bilan belgilandi. Fag bilan zararlangan bakterial hujayrada faqat radioaktiv P ni oʻz ichiga olgan fag zarrachalari hosil boʻlgan.
DNK va RNK molekulalarining tuzilishi va vazifalari.
Nuklein kislotalar tartibsiz tuzilishga ega biopolimerlar bo'lib, ularning monomerlari nukleotidlardir. Nukleotid uchta moddaning qoldiqlaridan iborat: fosfor kislotasi, uglevod - pentoza, azotli asos. DNK nukleotidlarida uglevod dezoksiriboza, RNKda esa riboza mavjud. DNK ni tashkil etuvchi purin va pirimidin azotli asoslarning qoldiqlari adenin, guanin, sitozin, timindir. RNK molekulalarida adenin, guanin, sitozin va urasil mavjud.
Nukleotidlar bir nukleotidning fosfor kislotasi qoldig'i va boshqa nukleotidning uglevodlari orqali bir-biriga "kislorod ko'prigi" deb ataladigan kuchli kovalent efir bog'i orqali bog'lanadi. Bog'lanish bir nukleotid uglevodining 5-uglerod atomi orqali boshqa nukleotid uglevodining 3-uglerod atomiga o'tadi. Nukleotidlar ketma-ketligi nuklein kislotalarning birlamchi tuzilishini ifodalaydi. RNK bitta polinukleotid zanjiridir. DNKning tuzilishi spiral shaklida o'ralgan qo'sh polinukleotid zanjiridir.
DNKning ikkilamchi tuzilishi birinchisiga nisbatan komplementarlik printsipiga ko'ra joylashtirilgan ikkinchi DNK zanjiri paydo bo'lganda hosil bo'ladi. Ikkinchi zanjir birinchisiga qarama-qarshi (antiparallel). Azotli asoslar molekula tekisligiga perpendikulyar tekislikda yotadi - bu spiral zinapoyaga o'xshaydi. Ushbu zinapoyaning panjaralari fosforik kislota qoldiqlari va uglevodlar, qadamlar esa azotli asoslardir.
Qarama-qarshi yo'naltirilgan zanjirlardagi har bir nukleotidni tashkil etuvchi azotli asoslar bir-biri bilan to'ldiruvchi vodorod bog'larini hosil qilish qobiliyatiga ega (har bir azotli asosning tuzilishidagi mavjud funktsional guruhlar tufayli). Adenil nukleotid timinga, guanil sitozinga va aksincha. O'z-o'zidan bu aloqalar mo'rt, ammo butun uzunligi bo'ylab qayta-qayta "tikilgan" DNK molekulasi juda kuchli aloqani ifodalaydi.
To'ldiruvchilik- bu azotli asoslarning bir-biriga fazoviy-strukturaviy va kimyoviy muvofiqligi; ular bir-biriga "qulf kaliti kabi" mos keladi.
Bitta DNK molekulasida 10 8 yoki undan ortiq nukleotid bo'lishi mumkin.
DNK molekulasining qo'sh antiparallel spiral sifatida tuzilishi 1953 yilda amerikalik biolog Jeyms Uotson va ingliz fizigi Frensis Krik tomonidan taklif qilingan.
Sayyoradagi har qanday tirik organizmning DNK molekulasi faqat to'rt turdagi nukleotidlardan iborat bo'lib, ular tarkibidagi azotli asoslarda bir-biridan farq qiladi: adenil, guanil, timin va sitozin. Unda ko'p qirralilik DNK. Ularning ketma-ketligi boshqacha, soni esa cheksizdir.
Tirik organizmlarning har bir turi uchun va har bir organizm uchun alohida, bu ketma-ketlik individual va qat'iydir. xos .
O'ziga xoslik DNKning tuzilishi shundan iboratki, molekulaning kimyoviy faol bo'limlari - azotli asoslar spiralning markaziga botiriladi va bir-biri bilan qo'shimcha aloqalar hosil qiladi, dezoksiriboza va fosfor kislotasi qoldiqlari esa periferiyada joylashgan va azotli asoslarga kirishni qoplaydi. - ular kimyoviy jihatdan faol emas. Ushbu struktura uzoq vaqt davomida kimyoviy barqarorlikni saqlab turishi mumkin. Irsiy ma'lumotni saqlash uchun yana nima kerak? DNK tuzilishining aynan shu xususiyatlari uning genetik ma'lumotni kodlash va ko'paytirish qobiliyatini belgilaydi.
DNKning kuchli tuzilishini yo'q qilish juda qiyin. Shunga qaramay, bu hujayrada muntazam ravishda sodir bo'ladi - RNK sintezi va hujayra bo'linishidan oldin DNK molekulasining ikki baravar ko'payishi.
Duplikatsiya, DNK replikatsiyasi maxsus ferment - DNK polimeraza qo'sh spiralni yechib, uni alohida iplarga ajratishidan boshlanadi - reduplikatsiya vilkasi hosil bo'ladi. Ferment fermuardagi qulf kabi ishlaydi. Har bir bir ipli zanjirda - reduplikatsiya vilkasining yopishqoq uchlarida - komplementarlik printsipiga ko'ra karioplazmadagi erkin nukleotidlardan yangi zanjir sintezlanadi. Ikki yangi DNK molekulasida bir zanjir asl ona zanjiri, ikkinchisi esa yangi qiz zanjiri bo'lib qoladi. Natijada, bitta DNK molekulasi o'rniga, asl nusxadagi kabi aniq nukleotid tarkibiga ega ikkita molekula paydo bo'ladi.
Tirik tizimlarda biz jonsiz tabiatda noma'lum bo'lgan reaktsiyalarning yangi turiga duch kelamiz. Ular chaqiriladi matritsa sintez reaktsiyalari . Matritsa sintezi matritsaga quyishga o'xshaydi: yangi molekulalar mavjud molekulalar tuzilishida belgilangan rejaga muvofiq sintezlanadi. Bu reaksiyalar sintez qilingan polimerlardagi monomer birliklarining aniq ketma-ketligini ta'minlaydi. Monomerlar reaktsiya sodir bo'ladigan matritsa bo'lib xizmat qiladigan molekulalarning ma'lum bir joyiga keladi. Agar bunday reaktsiyalar molekulalarning tasodifiy to'qnashuvi natijasida sodir bo'lsa, ular cheksiz sekinlik bilan boradi. Shablon prinsipiga asoslangan murakkab molekulalarning sintezi fermentlar yordamida tez va aniq amalga oshiriladi. Shablon sintezi nuklein kislotalar va oqsillar sintezidagi eng muhim reaksiyalar asosida yotadi. Hujayradagi matritsaning rolini nuklein kislotalar DNK yoki RNK molekulalari bajaradi. Polimer sintez qilinadigan monomerik molekulalar - nukleotidlar yoki aminokislotalar - komplementarlik printsipiga muvofiq, matritsada qat'iy belgilangan tartibda joylashadi va mahkamlanadi. Keyin monomer birliklari polimer zanjiriga ulanadi va tayyor polimer matritsani tark etadi. Shundan so'ng, matritsa yangi, aynan bir xil polimer molekulasini yig'ishga tayyor.
Matritsa tipidagi reaktsiyalar tirik hujayraning o'ziga xos xususiyatidir. Ular barcha tirik mavjudotlarning asosiy xususiyati - o'z turlarini ko'paytirish qobiliyatining asosidir.
Nuklein kislotalarning vazifalari– irsiy axborotni saqlash va uzatish. DNK molekulalari oqsilning asosiy tuzilishi haqidagi ma'lumotlarni kodlaydi. mRNK molekulalarining sintezi DNK matritsasida sodir bo'ladi. Bu jarayon "transkripsiya" deb ataladi. I-RNK "tarjima" jarayonida ma'lumotni oqsil molekulasidagi aminokislotalar ketma-ketligi ko'rinishida amalga oshiradi.
Har bir hujayraning DNKsi nafaqat hujayraning shaklini belgilovchi strukturaviy oqsillar, balki barcha ferment oqsillari, gormon oqsillari va boshqa oqsillar, shuningdek, RNKning barcha turlarining tuzilishi haqida ham ma'lumotlarni o'z ichiga oladi.
Ehtimol, nuklein kislotalar biologik xotiraning har xil turlarini - immunologik, nevrologik va boshqalarni ta'minlaydi, shuningdek, biosintetik jarayonlarni tartibga solishda muhim rol o'ynaydi.
Tegishli ma'lumotlar.
DNKning organizmlarda irsiy ma'lumot tashuvchisi sifatidagi rolining birinchi isboti nuklein kislotalarni o'rganishga katta e'tibor qaratdi. 1869 yilda F.Misher hujayra yadrolaridan maxsus moddani ajratib oldi va uni nuklein deb ataydi. 20 yildan keyin bu nom atama bilan almashtirildi nuklein kislotasi. 1924 yilda R. Felgen nuklein kislotalarni o'ziga xos bo'yash orqali sitologik aniqlash usulini ishlab chiqdi va DNK hujayralar yadrolarida, RNK esa sitoplazmada lokalizatsiya qilinishini ko'rsatdi. 1936 yilda A.N. Belozerskiy va I.I. Dubrovskaya o'simlik hujayralarining yadrolaridan sof shaklda DNKni ajratib oldi. 1930-yillarning boshlariga kelib. Nuklein kislota qandlari tuzilishining asosiy kimyoviy tamoyillari yoritib berildi va 1953 yilda DNKning strukturaviy modeli yaratildi.
Nuklein kislotalarning asosiy tuzilish birligi nukleotid, u kovalent bog'lar bilan bog'langan uchta kimyoviy jihatdan turli qismlardan iborat (5.2-rasm).
Guruch. 5.2. Strukturaviy formulalar: A- nukleotidlar; b- DNK; V - RNK (shuningdek qarang. 110-bet)

Guruch. 5.2. Tugatish. Strukturaviy formulalar: A- nukleotidlar; 6 - DNK; V- RNK
Birinchi qism beshta uglerod atomini o'z ichiga olgan shakar: deoksiriboza DNKda va riboza RNKda.
Nukleotidning ikkinchi qismi, purin yoki pirimidin azotli asos, shakarning birinchi uglerod atomi bilan kovalent bog'langan bo'lib, shunday tuzilishni hosil qiladi. nukleozid. DNK tarkibida purin asoslari mavjud - adenin(A) va guanin(D) - va pirimidin asoslari - timin(T) va sitozin(C). Tegishli nukleozidlar deoksiadenozin, deoksiguanozin, deoksitimidin va deoksitsitidin deb ataladi. RNK tarkibida pirimidin asosi bo'lgan DNK bilan bir xil purin asoslari mavjud sitozin, va timin o'rniga tarkibida mavjud urasil(U); tegishli nukleozidlar adenozin, guanozin, uridin va sitidin deb ataladi.
Nukleotidning uchinchi qismi fosfat guruhi bo'lib, u qo'shni nukleozidlarni bir shakarning 5-uglerod atomi va boshqasining 3-uglerod atomi o'rtasidagi fosfodiester bog'lari orqali polimer zanjiriga bog'laydi (5.2-rasm, b). V). Nukleotidlar shakarning 3" yoki 5 uglerod atomlariga ester bog'lari bilan bog'langan bir yoki bir nechta fosfat guruhlari bilan nukleozidlar deb ataladi. Nukleotidlarning sintezi nuklein kislotalarning sintezidan oldin sodir bo'ladi; shunga ko'ra, nukleotidlar nuklein kislotalarning kimyoviy yoki fermentativ gidrolizi mahsulotlaridir.
Nuklein kislotalar 5 va 3'-fosfodiester bog'lari bilan bog'langan mononukleotidlardan tashkil topgan juda uzun polimer zanjirlardir. Buzilmagan DNK molekulasida organizm turiga qarab bir necha mingdan ko'p milliongacha nukleotidlar, buzilmagan RNK molekulasi - 100 dan 100 minggacha yoki undan ko'p nukleotidlar mavjud.
E.Chargaffning turli xil shakllardagi DNK nukleotid tarkibini tahlil qilish natijalari turli azotli asoslar - adenin, guanin, timin, sitozinning molekulyar nisbati juda xilma-xil ekanligini ko'rsatdi. Shunday qilib, DNK 40-yillarda taxmin qilinganidek, bir xil tetranukleotidlardan tashkil topgan monoton polimer emasligi isbotlandi. XX asr va u nukleotid asoslarining ma'lum bir ketma-ketligi shaklida irsiy ma'lumotni saqlash va uzatish uchun zarur bo'lgan murakkablikka to'liq ega.
E. Chargaff tomonidan olib borilgan tadqiqotlar barcha DNK molekulalariga xos xususiyatni ham aniqladi: adeninning molyar miqdori timin tarkibiga, guaninning molyar miqdori sitozin tarkibiga teng. Bu tengliklar Chargaffning ekvivalentlik qoidasi deyiladi: [A] = [T], [G] = [C]; purinlar soni pirimidinlar soniga teng. Turlarga qarab faqat ([A] + [T])/([G] + [C]) nisbati o'zgaradi (5.1-jadval).
|
Bazalarning tarkibi |
Munosabat |
Asimmetriya |
|||||
|
sabablar |
|||||||
|
(A + T)/(G + C) |
|||||||
|
Hayvonlar |
|||||||
|
Toshbaqa |
|||||||
|
dengiz qisqichbaqasi |
|||||||
|
Dengiz tipratikoni |
|||||||
|
O'simliklar, qo'ziqorinlar |
|||||||
|
bug'doy urug'i |
|||||||
|
Qo'ziqorin Aspergillus niger |
|||||||
|
Bakteriyalar |
|||||||
|
Escherichia coli |
|||||||
|
Staphylococcus aureus |
|||||||
|
Clostridium perfringens |
|||||||
|
Brusela abortus |
|||||||
|
Sarcina lutea |
|||||||
|
Bakteriofaglar |
|||||||
|
FH 174 (virusli shakl) |
|||||||
|
FH 174 (replikativ shakl) |
|||||||
Asoslarning nisbati deyiladi nukleotid koeffitsienti(turlar) o'ziga xoslik. Chargaffning kashfiyoti DNKning muhim strukturaviy xususiyatini shakllantirdi, keyinchalik bu DNKning strukturaviy modelida J. Uotson va F. Krik (1953) tomonidan aks ettirildi, u aslida Chargaff qoidalari kombinatsiyalarning mumkin bo'lgan soniga hech qanday cheklovlar qo'ymasligini ko'rsatdi. DNK molekulalarini shakllantirishga qodir bo'lgan turli xil bazaviy ketma-ketliklar.
Nukleotidlarning o'ziga xosligi tushunchasi biologiyaning yangi bo'limining asosini tashkil etdi - gen sistematikasi, bu organizmlarning tabiiy tizimini qurish uchun nuklein kislotalarning tarkibi va tuzilishini solishtirish orqali ishlaydi.
Uotson-Krik modeliga ko'ra, DNK molekulasi komplementar printsipga ko'ra azotli asoslar orasidagi ko'ndalang vodorod bog'lari yordamida bir-biriga bog'langan ikkita polinukleotid zanjiridan (torlar, iplar) iborat (bir zanjirning adenini ikkita vodorod bog'i bilan bog'langan). qarama-qarshi zanjirning timin va turli zanjirli guanin va sitozin bir-biriga uchta vodorod bog'i bilan bog'langan). Bunda bir molekulaning ikkita polinukleotid zanjiri antiparallel bo'ladi, ya'ni bir zanjirning 3" uchi qarshisida ikkinchi zanjirning 5" uchi va aksincha (5.3-rasm). Biroq, zamonaviy ma'lumotlarni yodda tutish kerakki, ba'zi viruslarning genetik materiali bir zanjirli (bir zanjirli) DNK molekulalari bilan ifodalanadi. J. Uotson va F. Krik DNKning rentgen diffraksion tahliliga asoslanib, shuningdek, uning ikki zanjirli molekulasi chapdan o'ngga yo'nalishda buralib ketgan spiral shaklida ikkilamchi tuzilishga ega degan xulosaga kelishdi, keyinchalik u 5 deb nomlandi. -shakl (5.4-rasm). Bugungi kunga kelib, eng keng tarqalgan 5-shaklga qo'shimcha ravishda, boshqa konfiguratsiyaga ega bo'lgan DNK bo'limlarini - o'ng qo'lda (shakllar) aniqlash mumkinligi isbotlangan. A, C) va o'ngdan chapga o'ralgan (chapga o'ralgan yoki Z-shakli) (5.4-rasm). Ikkilamchi DNK strukturasining bu shakllari o'rtasida ma'lum farqlar mavjud (5.2-jadval). Masalan, nanometrlarda (nm) ifodalangan ikki torli spiraldagi ikki qo'shni azotli asoslar juftligi orasidagi masofa 5-forma va Z-shakl uchun turli qiymatlar bilan tavsiflanadi (mos ravishda 0,34 va 0,38 nm). . Shaklda. 5.5-rasmda DNKning "chap qo'l" va "o'ng qo'l" shakllarining zamonaviy uch o'lchovli modellari ko'rsatilgan.

Guruch. 5.3. Ikki zanjirli DNK molekulasining birlamchi strukturasining sxematik tasviri: A - adenin; G - guanin; T - timin; C - sitozin

Guruch. 5.4.

5.2-jadval
DNK juft spirallarining turli shakllarining xossalari
RNK molekulalari strukturaviy va funksional xususiyatlariga ko'ra bir necha turlarga bo'linadi: xabarchi RNK (mRNK yoki mRNK), ribosoma RNK (rRNK), transfer RNK (tRNK), kichik yadro RNK (snRNK) va boshqalar. Bundan farqli o'laroq. DNK, RNK molekulalari doimo bir zanjirli (bir zanjirli). Biroq, ular bir-birini to'ldiruvchi azotli asoslarning (A-U va G-C) o'zaro ta'siriga asoslangan bunday zanjirning alohida bo'limlarining bir-birini to'ldiruvchi ulanishi tufayli yanada murakkab (ikkilamchi) konfiguratsiyalarni hosil qilishi mumkin. Misol tariqasida, fenilalanin o'tkazuvchi RNK molekulasi uchun yonca bargi konfiguratsiyasini ko'rib chiqing (5.6-rasm).

Guruch. 5.6.
1953-yilda D.Uotson va F.Krik DNK tuzilishining modelini taklif qildilar, bu model quyidagi postulatlarga asoslanadi:
- 1. DNK 3" va 5" fosfodiester bog'lari bilan bog'langan nukleotidlardan tashkil topgan polimerdir.
- 2. DNK nukleotidlarining tarkibi Chargaff qoidalariga bo'ysunadi.
- 3. DNK molekulasi spiral zinapoyani eslatuvchi qo'sh spiralli tuzilishga ega bo'lib, buni birinchi marta M. Uilkins va R. Franklin tomonidan olingan DNK zanjirlarining rentgen difraksiya naqshlari tasdiqlaydi.
- 4. Nativ (tabiiy) DNK ni kislota-asos titrlashda ko'rsatilganidek, polimerning tuzilishi vodorod bog'lari bilan barqarorlashadi. Mahalliy DNKni titrlash va qizdirish uning fizik xususiyatlarini, xususan, yopishqoqligini sezilarli darajada o'zgartiradi, uni denatüratsiyalangan shaklga aylantiradi va kovalent aloqalar buzilmaydi.
Nuklein kislotalar mononukleotidlardan tashkil topgan yuqori molekulyar moddalar bo'lib, ular bir-biri bilan polimer zanjirida 3", 5" fosfodiefir bog'lari yordamida bog'langan va hujayralarga ma'lum tarzda qadoqlangan.
Nuklein kislotalar ikki turdagi biopolimerlar: ribonuklein kislotasi (RNK) va deoksiribonuklein kislotasi (DNK). Har bir biopolimer uglevod qoldig'i (riboza, dezoksiriboza) va azotli asoslardan biri (urasil, timin) bilan farq qiluvchi nukleotidlardan iborat. Ushbu farqlarga ko'ra, nuklein kislotalar o'z nomini oldi.
Dezoksiribonuklein kislotaning tuzilishi
Nuklein kislotalar birlamchi, ikkilamchi va uchinchi darajali tuzilishga ega.
DNKning birlamchi tuzilishi
DNKning asosiy tuzilishi chiziqli polinukleotid zanjiri bo'lib, unda mononukleotidlar 3 "5" fosfodiester bog'lari bilan bog'langan. Hujayrada nuklein kislota zanjirini yig'ish uchun boshlang'ich material 5"-trifosfat nukleozid bo'lib, u b va g fosforik kislota qoldiqlarini olib tashlash natijasida boshqa nukleozidning 3" uglerod atomini biriktirishga qodir. . Shunday qilib, bitta deoksiribozaning 3" uglerod atomi boshqa dezoksiribozaning 5" uglerod atomiga bitta fosfor kislotasi qoldig'i orqali kovalent bog'lanadi va nuklein kislotaning chiziqli polinukleotid zanjirini hosil qiladi. Shuning uchun nomi: 3", 5" fosfodiester bog'lari. Azotli asoslar bir zanjirning nukleotidlarini ulashda qatnashmaydi (1.-rasm).
Bir nukleotidning fosfor kislotasi molekulasi qoldig'i va boshqasining uglevodlari o'rtasidagi bunday bog'lanish polinukleotid molekulasining pentoza-fosfat skeletining hosil bo'lishiga olib keladi, bunda azotli asoslar birin-ketin yon tomonga biriktiriladi. Ularning nuklein kislota molekulalarining zanjirlarida joylashish ketma-ketligi qat'iy ravishda turli organizmlarning hujayralari uchun xosdir, ya'ni. o'ziga xos xususiyatga ega (Chargaff qoidasi).
Uzunligi zanjirdagi nukleotidlar soniga bog'liq bo'lgan chiziqli DNK zanjirining ikkita uchi bor: biri 3" uchi deb ataladi va erkin gidroksilni o'z ichiga oladi, ikkinchisi esa 5" uchi deb ataladi va fosforni o'z ichiga oladi. kislota qoldig'i. Sxema qutbli bo'lib, yo'nalishi 5"->3" va 3"->5" bo'lishi mumkin. Istisno - dumaloq DNK.
DNKning genetik "matn"i kod "so'zlari" - kodonlar deb ataladigan nukleotidlarning tripletlaridan iborat. Barcha turdagi RNKlarning birlamchi tuzilishi haqidagi ma'lumotlarni o'z ichiga olgan DNK bo'limlari strukturaviy genlar deb ataladi.
Polinukleotid DNK zanjirlari ulkan o'lchamlarga etadi, shuning uchun ular hujayrada ma'lum bir tarzda qadoqlanadi.
Chargaff (1949) DNK tarkibini o'rganar ekan, alohida DNK asoslari tarkibiga oid muhim naqshlarni aniqladi. Ular DNKning ikkilamchi tuzilishini ochishga yordam berdi. Bu naqshlar Chargaff qoidalari deb ataladi. Chargaff qoidalari
Ushbu qoidalar shuni ko'rsatadiki, DNKni qurishda purin va pirimidin asoslari emas, balki adenin bilan timin va guanin bilan sitozin juda qattiq mos kelishi (juftligi) kuzatilishi kerak. Ushbu qoidalarga asoslanib, 1953 yilda Uotson va Krik DNKning ikkilamchi strukturasining qo'sh spiral deb nomlangan modelini taklif qildilar (rasm). |

DNKning ikkilamchi tuzilishi
DNKning ikkilamchi strukturasi qoʻsh spiral boʻlib, uning modeli 1953-yilda D.Uotson va F.Krik tomonidan taklif qilingan.
DNK modelini yaratish uchun zarur shartlar
Dastlabki tahlillar natijasida har qanday kelib chiqadigan DNKda barcha to'rtta nukleotidlar teng molyar miqdorda borligiga ishonishdi. Biroq 1940-yillarda E.Chargaff va uning hamkasblari turli organizmlardan ajratilgan DNKni tahlil qilish natijasida ular tarkibida turli miqdoriy nisbatlarda azotli asoslar borligini aniq ko‘rsatdi. Chargaff shuni aniqladiki, bu nisbatlar bir xil turdagi organizmlarning barcha hujayralarining DNKlari uchun bir xil bo'lsa-da, har xil turdagi DNKlar ma'lum nukleotidlarning tarkibida sezilarli darajada farq qilishi mumkin. Bu azotli asoslar nisbatidagi farqlar qandaydir biologik kod bilan bog'liq bo'lishi mumkinligini ko'rsatdi. Turli DNK namunalarida individual purin va pirimidin asoslarining nisbati har xil bo'lsa-da, sinov natijalarini solishtirganda ma'lum bir naqsh paydo bo'ldi: barcha namunalarda purinlarning umumiy soni pirimidinlarning umumiy soniga teng edi (A +). G = T + C), adenin miqdori timin miqdoriga (A = T), guanin miqdori esa sitozin miqdoriga (G = C) teng edi. Sutemizuvchilar hujayralaridan ajratilgan DNK odatda adenin va timinga boyroq, guanin va sitozinga nisbatan kambag'al edi, bakteriyalar DNKsi esa guanin va sitozinga boyroq, adenin va timinga nisbatan kambag'al edi. Ushbu ma'lumotlar faktik materialning muhim qismini tashkil etdi, uning asosida keyinchalik DNK tuzilishining Uotson-Krik modeli qurilgan.
DNKning mumkin bo'lgan tuzilishining yana bir muhim bilvosita belgisi L. Paulingning oqsil molekulalarining tuzilishi haqidagi ma'lumotlari bilan ta'minlandi. Pauling oqsil molekulasidagi aminokislotalar zanjirining turli xil barqaror konfiguratsiyalari mumkinligini ko'rsatdi. Umumiy peptid zanjiri konfiguratsiyasi, a-spiral, muntazam spiral strukturadir. Ushbu tuzilish bilan zanjirning qo'shni burilishlarida joylashgan aminokislotalar o'rtasida vodorod aloqalarining shakllanishi mumkin. Pauling 1950 yilda polipeptid zanjirining a-spiral konfiguratsiyasini tasvirlab berdi va DNK molekulalari, ehtimol, vodorod bog'lari bilan tutilgan spiral tuzilishga ega bo'lishini taklif qildi.
Biroq, DNK molekulasining tuzilishi haqidagi eng qimmatli ma'lumotlar rentgen nurlari diffraktsiyasi tahlili natijalari bilan ta'minlandi. DNK kristalidan o'tuvchi rentgen nurlari difraksiyaga uchraydi, ya'ni ular ma'lum bir yo'nalishda burilib ketadi. Nurlarning burilish darajasi va tabiati molekulalarning tuzilishiga bog'liq. Rentgen nurlari diffraktsiyasi (3-rasm) tajribali ko'zga o'rganilayotgan moddaning molekulalarining tuzilishiga oid bir qator bilvosita ko'rsatkichlarni beradi. DNKning rentgen nurlanishining diffraktsiya naqshlarini tahlil qilish, azotli asoslar (tekis shaklga ega) plitalar to'plami kabi joylashtirilgan degan xulosaga keldi. Rentgen nurlari diffraktsiyasi kristalli DNK tuzilishining uchta asosiy davrini aniqladi: 0,34, 2 va 3,4 nm.

Watson-Crick DNK modeli
Chargaffning analitik ma'lumotlari, Uilkinsning rentgen naqshlari va molekuladagi atomlar orasidagi aniq masofalar, berilgan atomning bog'lanishlari orasidagi burchaklar va atomlarning o'lchamlari haqida ma'lumot bergan kimyogarlarning tadqiqotlari asosida Uotson va Krik DNK molekulasining alohida tarkibiy qismlarining fizik modellarini ma'lum miqyosda qurishni va ularni bir-biriga "sozlash" ni, natijada olingan tizim turli xil eksperimental ma'lumotlarga mos kelishini boshladi. [ko'rsatish] .
DNK zanjiridagi qo'shni nukleotidlar bir nukleotidning 5"-uglerod dezoksiriboza atomini keyingi nukleotidning 3"-uglerod dezoksiriboza atomi bilan bog'lab, fosfodiester ko'priklar orqali bog'langanligi avvalroq ma'lum edi. Uotson va Krik 0,34 nm davri DNK zanjiridagi ketma-ket nukleotidlar orasidagi masofaga mos kelishiga shubha qilishmagan. Bundan tashqari, 2 nm davri zanjirning qalinligiga to'g'ri keladi deb taxmin qilish mumkin. Va 3,4 nm davri qanday haqiqiy tuzilishga mos kelishini tushuntirish uchun Uotson va Krik, shuningdek, avvalroq Pauling zanjirning spiral shaklida o'ralganligini (yoki aniqrog'i, spiral chiziq hosil qiladi, chunki) bu so'zlarning qat'iy ma'nosida spiral bo'laklar kosmosda silindrsimon emas, balki konusning sirtini hosil qilganda olinadi). Keyin 3,4 nm davr bu spiralning ketma-ket burilishlari orasidagi masofaga to'g'ri keladi. Bunday spiral juda zich yoki biroz cho'zilgan bo'lishi mumkin, ya'ni uning burilishlari tekis yoki tik bo'lishi mumkin. 3,4 nm davri ketma-ket nukleotidlar orasidagi masofadan (0,34 nm) roppa-rosa 10 barobar ko'p bo'lganligi sababli, spiralning har bir to'liq aylanishida 10 ta nukleotid borligi aniq. Ushbu ma'lumotlardan Uotson va Krik diametri 2 nm, burilishlar orasidagi masofa 3,4 nm bo'lgan spiralga o'ralgan polinukleotid zanjirining zichligini hisoblashga muvaffaq bo'ldi. Ma'lum bo'lishicha, bunday zanjir allaqachon ma'lum bo'lgan DNKning haqiqiy zichligining yarmiga teng bo'lgan zichlikka ega bo'ladi. Men DNK molekulasi ikkita zanjirdan iborat deb taxmin qilishim kerak edi - bu nukleotidlarning qo'sh spiralidir.
Keyingi vazifa, albatta, qo'sh spiralni tashkil etuvchi ikkita zanjir o'rtasidagi fazoviy munosabatlarni aniqlashtirish edi. Uotson va Krik o'zlarining jismoniy modeli bo'yicha zanjirlarni joylashtirishning bir qancha variantlarini sinab ko'rib, barcha mavjud ma'lumotlar ikkita polinukleotid spirallari qarama-qarshi yo'nalishda harakatlanadigan variantga eng mos kelishini aniqladilar; bu holda shakar va fosfat qoldiqlaridan tashkil topgan zanjirlar qo'sh spiralning sirtini hosil qiladi va purinlar va pirimidinlar ichida joylashgan. Ikki zanjirga mansub, bir-biriga qarama-qarshi joylashgan asoslar vodorod bog'lari orqali juft bo'lib bog'langan; Aynan shu vodorod aloqalari zanjirlarni bir-biriga bog'lab turadi va shu bilan molekulaning umumiy konfiguratsiyasini o'rnatadi.
DNKning qo'sh spiralini zinapoyalari gorizontal holatda bo'lishi uchun spiral tarzda buralib ketgan arqon narvon sifatida tasavvur qilish mumkin. Keyin ikkita uzunlamasına arqon shakar va fosfat qoldiqlari zanjirlariga mos keladi va shpallar vodorod aloqalari bilan bog'langan azotli asoslarning juftlariga mos keladi.
Mumkin bo'lgan modellarni keyingi o'rganish natijasida Uotson va Krik har bir "shpal" bitta purin va bitta pirimidindan iborat bo'lishi kerak degan xulosaga kelishdi; 2 nm (qo'sh spiral diametriga to'g'ri keladigan) davrda ikkita purin uchun etarli joy bo'lmaydi va ikkita pirimidin to'g'ri vodorod aloqalarini hosil qilish uchun bir-biriga etarlicha yaqin bo'lolmaydi. Batafsil modelni chuqur o'rganish shuni ko'rsatdiki, adenin va sitozin mos o'lchamdagi birikma hosil qilganda, ular o'rtasida vodorod aloqalari hosil bo'ladigan tarzda joylashishi mumkin emas. Shunga o'xshash hisobotlar guanin - timin birikmasini chiqarib tashlashga majbur qildi, adenin - timin va guanin - sitozin birikmalari esa juda maqbul bo'lib chiqdi. Vodorod aloqalarining tabiati shundayki, adenin timin bilan, guanin esa sitozin bilan juft hosil qiladi. O'ziga xos asoslarni juftlashtirish g'oyasi "Chargaff qoidasi" ni tushuntirishga imkon berdi, unga ko'ra har qanday DNK molekulasida adenin miqdori doimo timin tarkibiga, guanin miqdori esa har doim miqdorga teng bo'ladi. sitozin. Adenin va timin o'rtasida ikkita, guanin va sitozin o'rtasida uchta vodorod aloqasi hosil bo'ladi. Ushbu o'ziga xoslik tufayli bir zanjirdagi har bir adeninga nisbatan vodorod aloqalarining shakllanishi boshqasida timin hosil bo'lishiga olib keladi; xuddi shunday, faqat sitozin har bir guaninga qarama-qarshi bo'lishi mumkin. Shunday qilib, zanjirlar bir-birini to'ldiradi, ya'ni bir zanjirdagi nukleotidlar ketma-ketligi ularning ikkinchisida ketma-ketligini o'ziga xos tarzda belgilaydi. Ikki zanjir qarama-qarshi yo'nalishda harakat qiladi va ularning terminal fosfat guruhlari qo'sh spiralning qarama-qarshi uchlarida joylashgan.
O'z tadqiqotlari natijasida 1953 yilda Uotson va Krik DNK molekulasi strukturasi modelini taklif qildilar (3-rasm), bu hozirgi kungacha dolzarbligicha qolmoqda. Modelga ko'ra, DNK molekulasi ikkita qo'shimcha polinukleotid zanjiridan iborat. Har bir DNK zanjiri bir necha o'n minglab nukleotidlardan tashkil topgan polinukleotiddir. Unda qo'shni nukleotidlar fosfor kislotasi qoldig'i va dezoksiribozaning kuchli kovalent bog' bilan bog'lanishi tufayli muntazam pentoza-fosfat magistralini hosil qiladi. Bir polinukleotid zanjirining azotli asoslari boshqasining azotli asoslariga qarama-qarshi qat'iy belgilangan tartibda joylashtirilgan. Polinukleotid zanjirida azotli asoslarning almashinishi tartibsizdir.
DNK zanjirida azotli asoslarning joylashuvi bir-birini to'ldiruvchi (yunoncha "to'ldiruvchi" - qo'shish), ya'ni. Timin (T) har doim adeninga (A), faqat sitozin (C) guaninga (G) qarshi. Bu A va T, shuningdek, G va C bir-biriga qat'iy mos kelishi bilan izohlanadi, ya'ni. bir-birini to‘ldiradi. Bu yozishmalar purin va pirimidin juftligida vodorod aloqalarini hosil qilish imkonini beruvchi asoslarning kimyoviy tuzilishi bilan belgilanadi. A va T o'rtasida ikkita, G va C o'rtasida uchta bog'lanish mavjud. Bu bog'lanishlar DNK molekulasining kosmosda qisman barqarorlashuvini ta'minlaydi. Ikki tomonlama spiralning barqarorligi A = T bog'lanishlariga nisbatan barqarorroq bo'lgan G≡C bog'lanishlari soniga to'g'ridan-to'g'ri proportsionaldir.
Bitta DNK zanjirida nukleotidlarning joylashishining ma'lum ketma-ketligi komplementarlik printsipiga ko'ra boshqa zanjirning nukleotidlarini o'rnatishga imkon beradi.
Bundan tashqari, suvli eritmada aromatik tuzilishga ega bo'lgan azotli asoslar bir-birining ustiga joylashib, go'yo tangalar to'plamini tashkil etishi aniqlandi. Organik molekulalarning steklarini hosil qilish jarayoni stacking deb ataladi. Ko'rib chiqilayotgan Uotson-Krik modelining DNK molekulasining polinukleotid zanjirlari o'xshash fizik-kimyoviy holatga ega, ularning azotli asoslari tangalar to'plami shaklida joylashgan bo'lib, ularning tekisliklari orasida van der Vaals o'zaro ta'siri (stacking o'zaro ta'sirlari) paydo bo'ladi.
Van-der-Vaals kuchlari (vertikal) taʼsirida polinukleotid zanjiridagi asoslar tekisliklari orasidagi komplementar asoslar (gorizontal) va stacking oʻzaro taʼsirlari oʻrtasidagi vodorod aloqalari DNK molekulasini kosmosda qoʻshimcha barqarorlashtirishni taʼminlaydi.
Ikkala zanjirning shakar fosfat umurtqalari tashqariga, asoslari esa ichkariga, bir-biriga qaragan. DNKdagi zanjirlarning yo'nalishi antiparallel (ulardan biri 5"->3" yo'nalishiga ega, ikkinchisi - 3"->5", ya'ni bir zanjirning 3" uchi 5" uchiga qarama-qarshi joylashgan. boshqa.). Zanjirlar umumiy o'qga ega bo'lgan o'ng qo'lli spirallarni hosil qiladi. Spiralning bir burilishi 10 nukleotid, burilish hajmi 3,4 nm, har bir nukleotidning balandligi 0,34 nm, spiral diametri 2,0 nm. Bir ipning ikkinchi ip atrofida aylanishi natijasida DNK qo'sh spiralining katta yivi (diametri taxminan 20 Å) va kichik yivi (diametri taxminan 12 Å) hosil bo'ladi. Uotson-Krik qo'sh spiralining bu shakli keyinchalik B-shakl deb nomlandi. Hujayralarda DNK odatda eng barqaror bo'lgan B shaklida mavjud.
DNKning funktsiyalari
Taklif etilayotgan model dezoksiribonuklein kislotasining ko'plab biologik xususiyatlarini, shu jumladan genetik ma'lumotni saqlashni va 4 nukleotidning turli xil ketma-ket birikmalari bilan ta'minlangan genlarning xilma-xilligini va genetik kodning mavjudligi, o'z-o'zini ko'paytirish qobiliyatini tushuntirdi. va replikatsiya jarayoni bilan ta'minlangan genetik ma'lumotni uzatish va genetik ma'lumotni oqsillar shaklida amalga oshirish, shuningdek, ferment oqsillari yordamida hosil bo'lgan har qanday boshqa birikmalar.
DNKning asosiy funktsiyalari.
- DNK genetik ma'lumotlarning tashuvchisi bo'lib, bu genetik kodning mavjudligi bilan ta'minlanadi.
- Hujayralar va organizmlarning avlodlari bo'ylab genetik ma'lumotlarni ko'paytirish va uzatish. Bu funksiya replikatsiya jarayoni bilan ta'minlanadi.
- Genetik ma'lumotni oqsillar, shuningdek, ferment oqsillari yordamida hosil bo'lgan boshqa birikmalar shaklida amalga oshirish. Bu funksiya transkripsiya va tarjima jarayonlari bilan ta'minlanadi.

Ikki zanjirli DNKni tashkil qilish shakllari
DNK bir necha turdagi qo'sh spiral hosil qilishi mumkin (4-rasm). Hozirgi vaqtda oltita shakl allaqachon ma'lum (A dan E va Z-shakl).
Rozalind Franklin aniqlaganidek, DNKning strukturaviy shakllari nuklein kislota molekulasining suv bilan toʻyinganligiga bogʻliq. DNK tolalarini rentgen difraksion tahlilidan foydalangan holda tadqiq qilishda rentgen tasviri tajribaning ushbu tolaning suv bilan toʻyinganligi darajasining nisbiy namligiga tubdan bogʻliqligi koʻrsatilgan. Agar tolalar suv bilan etarlicha to'yingan bo'lsa, unda bitta rentgenogramma olingan. Quritilganda, namlik yuqori bo'lgan tolaning rentgenogrammasidan juda farq qiladigan butunlay boshqa rentgen naqshlari paydo bo'ldi.
Yuqori namlikdagi DNK molekulasi B-formasi deb ataladi. Fiziologik sharoitda (tuzning past konsentratsiyasi, yuqori hidratsiya darajasi) DNKning dominant strukturaviy turi B-shakli (ikki zanjirli DNKning asosiy shakli - Uotson-Krik modeli). Bunday molekulaning spiral qadami 3,4 nm. Har bir navbatda 10 ta qo'shimcha juftlik "tangalar" - azotli asoslarning o'ralgan to'plamlari shaklida mavjud. Staklar ikkita qarama-qarshi "tangalar" orasidagi vodorod bog'lari bilan birlashtiriladi va o'ng qo'l spiraliga o'ralgan fosfodiester magistralining ikkita lentasi bilan "o'raladi". Azotli asoslar tekisliklari spiral o'qiga perpendikulyar. Qo'shni to'ldiruvchi juftliklar bir-biriga nisbatan 36 ° ga aylantiriladi. Spiralning diametri 20Å, purin nukleotidi 12Å va pirimidin nukleotidi 8Å ni egallaydi.
Pastroq namlikdagi DNK molekulasi A-formasi deb ataladi. A-shakli kamroq yuqori hidratsiya sharoitida va Na + yoki K + ionlarining yuqori miqdorida hosil bo'ladi. Ushbu kengroq o'ng qo'l spiral konformatsiyasi har bir burilishda 11 ta asosiy juftlikka ega. Azotli asoslar tekisliklari spiral o'qiga nisbatan kuchliroq moyillikka ega bo'lib, ular odatdagidan 20 ° ga og'ishgan. Bu diametri 5Å bo'lgan ichki bo'shliq mavjudligini nazarda tutadi. Qo'shni nukleotidlar orasidagi masofa 0,23 nm, burilish uzunligi 2,5 nm va spiralning diametri 2,3 nm.
DNKning A shakli dastlab unchalik muhim emas deb hisoblangan. Biroq, keyinchalik ma'lum bo'ldiki, DNKning A-shakli, xuddi B-shakli kabi, juda katta biologik ahamiyatga ega. Shablon-primer kompleksidagi RNK-DNK spiral A-shaklga, shuningdek, RNK-RNK spiral va RNK soch turmagi tuzilmalariga ega (ribozaning 2'-gidroksil guruhi RNK molekulalarining B-shaklini hosil qilishiga to'sqinlik qiladi). DNKning A-shakli sporalarda uchraydi. Aniqlanishicha, DNK ning A-shakli B shakliga qaraganda UV nurlariga 10 marta chidamli.
A-shakl va B-shakl DNKning kanonik shakllari deb ataladi.
C-E shakllari shuningdek, o'ng qo'llar, ularning shakllanishi faqat maxsus tajribalarda kuzatilishi mumkin va, aftidan, ular in vivo mavjud emas. DNK ning C shakli B DNK ga o'xshash tuzilishga ega. Har bir burilishda asosiy juftliklar soni 9,33, spiral burilish uzunligi 3,1 nm. Asosiy juftliklar o'qga perpendikulyar holatga nisbatan 8 graduslik burchak ostida moyil bo'ladi. Yivlar hajmi jihatidan B-DNK yivlariga o'xshaydi. Bunday holda, asosiy truba biroz sayozroq, kichik truba esa chuqurroqdir. Tabiiy va sintetik DNK polinukleotidlari C shakliga aylanishi mumkin.
| Jadval 1. DNK tuzilmalarining ayrim turlarining xarakteristikalari | |||
| Spiral turi | A | B | Z |
| Spiral qadam | 0,32 nm | 3,38 nm | 4,46 nm |
| Spiral burilish | To'g'ri | To'g'ri | Chapga |
| Har bir burilishda tayanch juftliklar soni | 11 | 10 | 12 |
| Asosiy tekisliklar orasidagi masofa | 0,256 nm | 0,338 nm | 0,371 nm |
| Glikozid bog'lanish konformatsiyasi | qarshi | qarshi | anti-C gunoh-G |
| Furanoz halqasining konformatsiyasi | C3"-endo | C2"-endo | C3"-endo-G C2"-endo-C |
| Yiv kengligi, kichik/katta | 1,11/0,22 nm | 0,57/1,17 nm | 0,2/0,88 nm |
| Chuqurlik chuqurligi, kichik/katta | 0,26/1,30 nm | 0,82/0,85 nm | 1,38/0,37 nm |
| Spiral diametri | 2,3 nm | 2,0 nm | 1,8 nm |
DNKning strukturaviy elementlari
(kanonik bo'lmagan DNK tuzilmalari)
DNKning strukturaviy elementlariga ba'zi maxsus ketma-ketliklar bilan cheklangan g'ayrioddiy tuzilmalar kiradi:
|

Z shaklidagi DNK 1979 yilda geksanukleotid d(CG)3 - ni o'rganayotganda kashf etilgan. Uni MIT professori Aleksandr Rich va uning hamkasblari kashf etgan. Z-shakli DNKning eng muhim tuzilmaviy elementlaridan biriga aylandi, chunki uning shakllanishi DNK mintaqalarida kuzatilgan, bu erda purinlar pirimidinlar bilan almashinadi (masalan, 5'-GCGCGC-3') yoki takroriy 5. Metillangan sitozinni o'z ichiga olgan "-CGCGCG-3". Z-DNKning shakllanishi va barqarorlashuvining muhim sharti uning tarkibidagi purin nukleotidlarining sin konformatsiyasida, antikonformatsiyada pirimidin asoslari bilan almashinishi edi.
Tabiiy DNK molekulalari, agar ular (CG) n kabi ketma-ketliklarni o'z ichiga olmasa, asosan o'ng qo'l B shaklida mavjud. Ammo, agar bunday ketma-ketliklar DNKning bir qismi bo'lsa, u holda bu bo'limlar, eritmaning ion kuchi yoki fosfodiester ramkasidagi manfiy zaryadni neytrallaydigan kationlar o'zgarganda, bu bo'limlar Z-shakliga, DNKning boshqa bo'limlari esa DNKga aylanishi mumkin. zanjir klassik B shaklida qoladi. Bunday o'tish imkoniyati shuni ko'rsatadiki, DNK qo'sh spiralidagi ikkita zanjir dinamik holatda bo'lib, o'ng qo'l shaklidan chap qo'lga va aksincha, bir-biriga nisbatan yechilishi mumkin. DNK strukturasining konformatsion o'zgarishlariga imkon beruvchi bunday labillikning biologik oqibatlari hali to'liq tushunilmagan. Z-DNK bo'limlari ma'lum genlarning ifodalanishini tartibga solishda ma'lum rol o'ynaydi va genetik rekombinatsiyada ishtirok etadi, deb ishoniladi.
DNKning Z-shakli chap qo'l qo'sh spiral bo'lib, unda fosfodiester magistral molekula o'qi bo'ylab zigzag shaklida joylashgan. Shuning uchun molekulaning nomi (zigzag)-DNK. Z-DNK tabiatda ma'lum bo'lgan eng kam burilgan (har bir burilishda 12 ta asosiy juft) va eng nozik DNKdir. Qo‘shni nukleotidlar orasidagi masofa 0,38 nm, burilish uzunligi 4,56 nm, Z-DNK diametri esa 1,8 nm. Bundan tashqari, bu DNK molekulasining ko'rinishi bitta yiv mavjudligi bilan ajralib turadi.
DNKning Z shakli prokaryotik va eukaryotik hujayralarda topilgan. Endi Z-shaklini DNKning B-shaklidan ajrata oladigan antikorlar olindi. Bu antikorlar Drosophila (Dr. melanogaster) tuprik bezi hujayralarining gigant xromosomalarining ma'lum hududlari bilan bog'lanadi. Ushbu xromosomalarning g'ayrioddiy tuzilishi tufayli bog'lanish reaktsiyasini kuzatish oson, ularda zichroq hududlar (disklar) kamroq zichroq hududlar (disklar) bilan farq qiladi. Z-DNK hududlari interdisklarda joylashgan. Bundan kelib chiqadiki, Z-shakli aslida tabiiy sharoitda mavjud, ammo Z-shaklning alohida bo'limlarining o'lchamlari hali ham noma'lum.

(invertorlar) DNKdagi eng mashhur va tez-tez uchraydigan asosiy ketma-ketliklardir. Palindrom - chapdan o'ngga va aksincha bir xil o'qiladigan so'z yoki ibora. Bunday so‘z yoki iboralarga misol bo‘la oladi: HUT, KAZAK, SEL, AZOR PANGASIGA TUSHGAN AZOR. DNK bo'limlariga qo'llanilganda, bu atama (palindrom) zanjir bo'ylab o'ngdan chapga va chapdan o'ngga ("kulba" so'zidagi harflar kabi) nukleotidlarning bir xil almashinuvini anglatadi.
Palindrom ikkita DNK zanjiriga nisbatan ikkinchi tartibli simmetriyaga ega bo'lgan asosiy ketma-ketliklarning teskari takrorlanishi mavjudligi bilan tavsiflanadi. Bunday ketma-ketliklar, aniq sabablarga ko'ra, o'z-o'zini to'ldiradi va soch turmagi yoki xoch shaklidagi tuzilmalarni shakllantirishga moyildir (rasm). Soch iplari tartibga soluvchi oqsillarga xromosoma DNKsining genetik matni qaerdan ko'chirilganligini aniqlashga yordam beradi.
Xuddi shu DNK zanjirida teskari takrorlanish mavjud bo'lsa, ketma-ketlik oyna takrori deb ataladi. Ko'zgularni takrorlash o'z-o'zini to'ldiruvchi xususiyatlarga ega emas va shuning uchun soch turmagi yoki xoch shaklidagi tuzilmalarni shakllantirishga qodir emas. Ushbu turdagi ketma-ketliklar deyarli barcha yirik DNK molekulalarida uchraydi va bir necha tayanch juftlikdan bir necha ming tayanch juftlikgacha bo'lishi mumkin.
Eukaryotik hujayralarda xochsimon tuzilmalar ko'rinishidagi palindromlarning mavjudligi isbotlanmagan, garchi ma'lum miqdordagi xochsimon tuzilmalar in vivo jonli ravishda E. coli hujayralarida aniqlangan. RNK yoki bir ipli DNKda o'z-o'zini to'ldiruvchi ketma-ketliklarning mavjudligi eritmalardagi nuklein kislota zanjirining ma'lum bir fazoviy tuzilishga katlanishining asosiy sababi bo'lib, ko'plab "soch iplari" shakllanishi bilan tavsiflanadi.

H shaklidagi DNK uch DNK zanjiridan hosil bo'lgan spiraldir - DNK uch spiral. Bu uchinchi bir torli DNK zanjiriga ega bo'lgan Uotson-Krik qo'sh spiral majmuasi bo'lib, uning asosiy yiviga kirib, Hoogsteen juftligini hosil qiladi.
Bunday tripleksning hosil bo'lishi DNK qo'sh spiralining shunday buklanishi natijasida sodir bo'ladi, uning kesimining yarmi qo'sh spiral shaklida qoladi, ikkinchi yarmi esa ajratiladi. Bunda uzilgan spirallardan biri qo`sh spiralning birinchi yarmi bilan yangi struktura - uchburchak spiralni hosil qiladi, ikkinchisi esa tuzilmagan bo`lib, bir ipli kesma shaklida chiqadi. Ushbu tizimli o'tishning o'ziga xos xususiyati uning protonlari yangi strukturani barqarorlashtiradigan muhitning pH ga keskin bog'liqligidir. Ushbu xususiyat tufayli yangi struktura DNKning H-shakli deb nomlandi, uning shakllanishi ko'zgu takrori bo'lgan gomopurin-gomopirimidin hududlarini o'z ichiga olgan supero'ralgan plazmidlarda topilgan.
Keyingi tadqiqotlarda ba'zi bir gomopurin-homopirimidin ikki ipli polinukleotidlarning tarkibiy o'tishini o'z ichiga olgan uch zanjirli tuzilma hosil qilgan holda amalga oshirish mumkinligi aniqlandi:
- bitta gomopurin va ikkita gomopirimidin zanjiri ( Py-Pu-Py tripleksi) [Hoogsteen shovqini].
Py-Pu-Py tripleksining tarkibiy bloklari kanonik izomorf CGC+ va TAT triadalaridir. Tripleksni barqarorlashtirish CGC+ triadasining protonatsiyasini talab qiladi, shuning uchun bu triplekslar eritmaning pH ga bog'liq.
- bitta gomopirimidin va ikkita gomopurin zanjiri ( Py-Pu-Pu tripleksi) [teskari Hoogsteen o'zaro ta'siri].
Py-Pu-Pu tripleksining tarkibiy bloklari kanonik izomorf CGG va TAA triadalaridir. Py-Pu-Pu triplekslarining muhim xususiyati ularning barqarorligining ikki marta zaryadlangan ionlar mavjudligiga bog'liqligi va turli ketma-ketlikdagi triplekslarni barqarorlashtirish uchun turli ionlar talab qilinadi. Py-Pu-Pu triplekslarining shakllanishi ularning tarkibiy nukleotidlarining protonatsiyasini talab qilmaganligi sababli, bunday triplekslar neytral pH da mavjud bo'lishi mumkin.
Eslatma: to'g'ridan-to'g'ri va teskari Hoogsteen o'zaro ta'siri 1-metiltiminning simmetriyasi bilan izohlanadi: 180 ° ga aylanish natijasida O2 atomi O4 atomi o'rnini egallaydi, vodorod aloqalari tizimi saqlanib qoladi.
Ikki turdagi uchburchaklar ma'lum:
- uchinchi ipning qutbliligi Watson-Crick dupleksining gomopurin zanjirining qutbliligiga to'g'ri keladigan parallel uchburchaklar.
- antiparallel uch spirallar, ularda uchinchi va gomopurin zanjirlarining qutblari qarama-qarshidir.

G-to'rtlik- 4 zanjirli DNK. Ushbu tuzilma to'rtta guanin bo'lsa, hosil bo'ladi, ular G-quadroplex deb ataladigan - to'rtta guaninning dumaloq raqsini tashkil qiladi.
Bunday tuzilmalarning paydo bo'lishi mumkinligi haqidagi birinchi maslahatlar Uotson va Krikning yutuq ishlaridan ancha oldin - 1910 yilda olingan. Keyin nemis kimyogari Ivar Bang DNK tarkibiy qismlaridan biri - guanosinik kislota - yuqori konsentratsiyalarda jellar hosil qilishini, DNKning boshqa komponentlarida esa bunday xususiyatga ega emasligini aniqladi.
1962 yilda rentgen nurlari diffraktsiya usulidan foydalanib, ushbu gelning hujayra tuzilishini o'rnatish mumkin edi. U to'rtta guanin qoldig'idan iborat bo'lib, bir-birini aylana shaklida bog'lab, xarakterli kvadrat hosil qiladi. Markazda bog'lanish metall ioni (Na, K, Mg) tomonidan quvvatlanadi. Xuddi shu tuzilmalar DNKda ko'p guanin bo'lsa, paydo bo'lishi mumkin. Ushbu tekis kvadratchalar (G-kvartetlar) juda barqaror, zich tuzilmalarni (G-to'rtlik) hosil qilish uchun yig'iladi.
DNKning to'rtta alohida zanjiri to'rt zanjirli komplekslarga to'qilishi mumkin, ammo bu istisno. Ko'pincha nuklein kislotaning bitta zanjiri oddiygina tugunga bog'lanib, xarakterli qalinlashuvlarni hosil qiladi (masalan, xromosomalarning uchlarida) yoki guaninga boy bo'lgan ba'zi bir mintaqada ikki ipli DNK mahalliy to'rtlik hosil qiladi.
Xromosomalarning uchlarida - telomerlarda va o'simta promotorlarida to'rtburchaklar mavjudligi eng ko'p o'rganilgan. Biroq, inson xromosomalarida bunday DNKning lokalizatsiyasining to'liq tasviri hali ham ma'lum emas.
Chiziqli shakldagi bu noodatiy DNK tuzilmalarining barchasi B shaklidagi DNKga nisbatan beqaror. Biroq, DNK ko'pincha topologik taranglikning dumaloq shaklida mavjud bo'lsa, unda supercoiling deb ataladi. Bunday sharoitda kanonik bo'lmagan DNK tuzilmalari osongina hosil bo'ladi: Z-shakllari, "xochlar" va "sochlar", H-shakllari, guanin kvadruplekslari va i-motiflar.
- Supero'ralgan shakl - pentoza fosfat magistraliga zarar bermasdan hujayra yadrosidan chiqarilganda qayd etiladi. U o'ta o'ralgan yopiq halqalar shakliga ega. Supero'ralgan holatda DNK qo'sh spiral kamida bir marta "o'z-o'zidan o'raladi", ya'ni u kamida bitta superburilishni o'z ichiga oladi (sakkizinchi raqam shaklini oladi).
- DNKning bo'shashgan holati - bitta uzilish (bir ipning uzilishi) bilan kuzatiladi. Bunday holda, superkoillar yo'qoladi va DNK yopiq halqa shaklini oladi.
- DNK ning chiziqli shakli qo'sh spiralning ikkita zanjiri uzilganda kuzatiladi.
DNKning uchinchi darajali tuzilishi
DNKning uchinchi darajali tuzilishi qo'sh spiralli molekulaning bo'shliqda qo'shimcha buralishi - uning o'ta burilishi natijasida hosil bo'ladi. Eukaryotik hujayralardagi DNK molekulasining superkoillanishi, prokariotlardan farqli o'laroq, oqsillar bilan komplekslar shaklida sodir bo'ladi.
Eukariotlarning deyarli barcha DNKlari yadrolarning xromosomalarida, faqat oz miqdori mitoxondriyalarda va o'simliklarda, plastidlarda mavjud. Eukaryotik hujayralar xromosomalarining asosiy moddasi (shu jumladan odam xromosomalari) xromatin bo'lib, ikki zanjirli DNK, giston va giston bo'lmagan oqsillardan iborat.
Giston kromatin oqsillari
Gistonlar oddiy oqsillar bo'lib, xromatinning 50% ni tashkil qiladi. Barcha o'rganilgan hayvon va o'simlik hujayralarida gistonlarning beshta asosiy klassi topildi: H1, H2A, H2B, H3, H4, hajmi, aminokislotalar tarkibi va zaryadi (har doim ijobiy) bo'yicha farqlanadi.
Sutemizuvchilar gistoni H1 taxminan 215 ta aminokislotani o'z ichiga olgan bitta polipeptid zanjiridan iborat; boshqa gistonlarning o'lchamlari 100 dan 135 aminokislotagacha o'zgarib turadi. Ularning barchasi spirallangan va diametri taxminan 2,5 nm bo'lgan globulaga o'ralgan va juda ko'p miqdorda musbat zaryadlangan aminokislotalar lizin va argininni o'z ichiga oladi. Gistonlar atsetillangan, metillangan, fosforlangan, poli(ADP)-ribosillangan bo'lishi mumkin, H2A va H2B gistonlari ubiquitin bilan kovalent bog'langan. Gistonlar tomonidan strukturaning shakllanishi va funktsiyalarini bajarishda bunday o'zgarishlarning roli hali to'liq o'rganilmagan. Bu ularning DNK bilan o'zaro ta'sir qilish qobiliyati va gen ta'sirini tartibga solish mexanizmlaridan birini ta'minlashi taxmin qilinadi.
Gistonlar DNK bilan birinchi navbatda DNK ning manfiy zaryadlangan fosfat guruhlari va gistonlarning musbat zaryadlangan lizin va arginin qoldiqlari oʻrtasida hosil boʻlgan ionli bogʻlar (tuz koʻprigi) orqali oʻzaro taʼsir qiladi.
Giston bo'lmagan xromatin oqsillari
Giston bo'lmagan oqsillar, gistonlardan farqli o'laroq, juda xilma-xildir. DNK bilan bog'langan giston bo'lmagan oqsillarning 590 tagacha turli fraktsiyalari ajratilgan. Ular kislotali oqsillar deb ham ataladi, chunki ularning tuzilishida kislotali aminokislotalar (ular polianionlar) ustunlik qiladi. Giston bo'lmagan oqsillarning xilma-xilligi xromatin faolligini o'ziga xos tartibga solish bilan bog'liq. Masalan, DNK replikatsiyasi va ekspressiyasi uchun zarur bo'lgan fermentlar xromatin bilan vaqtincha bog'lanishi mumkin. Boshqa oqsillar, aytaylik, turli xil tartibga solish jarayonlarida ishtirok etadiganlar, DNK bilan faqat ma'lum to'qimalarda yoki differentsiatsiyaning ma'lum bosqichlarida bog'lanadi. Har bir oqsil DNK nukleotidlarining ma'lum bir ketma-ketligiga (DNK sayti) qo'shimcha hisoblanadi. Bu guruhga quyidagilar kiradi:
- saytga xos sink barmoq oqsillari oilasi. Har bir "sink barmog'i" 5 ta nukleotid juftidan tashkil topgan ma'lum bir saytni taniydi.
- saytga xos oqsillar oilasi - homodimerlar. DNK bilan aloqada bo'lgan bunday oqsilning bo'lagi spiral-burilish-spiral tuzilishiga ega.
- yuqori harakatchanlikli gel oqsillari (HMG oqsillari) xromatin bilan doimo bog'langan tarkibiy va tartibga soluvchi oqsillar guruhidir. Ularning molekulyar og'irligi 30 kDa dan kam bo'lib, zaryadlangan aminokislotalarning yuqori miqdori bilan ajralib turadi. Past molekulyar og'irligi tufayli HMG oqsillari poliakrilamid gel elektroforez paytida yuqori harakatchanlikka ega.
- replikatsiya, transkripsiya va tuzatish fermentlari.
DNK va RNK sintezida ishtirok etadigan strukturaviy, tartibga soluvchi oqsillar va fermentlar ishtirokida nukleosoma ipi oqsillar va DNKning yuqori kondensatsiyalangan kompleksiga aylanadi. Olingan struktura dastlabki DNK molekulasidan 10 000 marta qisqa.

Xromatin
Xromatin yadro DNKsi va noorganik moddalarga ega bo'lgan oqsillar majmuasidir. Xromatinning asosiy qismi faol emas. U zich o'ralgan, kondensatsiyalangan DNKni o'z ichiga oladi. Bu heterokromatin. Konstitutsiyaviy, genetik jihatdan faol bo'lmagan xromatin (sun'iy yo'ldosh DNK) mavjud bo'lib, ular ifodalanmagan hududlardan iborat va fakultativ - bir qator avlodlarda faol bo'lmagan, ammo ma'lum sharoitlarda ifoda etishga qodir.
Faol kromatin (euchromatin) kondensatsiyalanmagan, ya'ni. kamroq mahkamlangan. Turli hujayralarda uning miqdori 2 dan 11% gacha. Miya hujayralarida u eng ko'p - 10-11%, jigar hujayralarida - 3-4 va buyrak hujayralarida - 2-3%. Euchromatinning faol transkripsiyasi qayd etilgan. Bundan tashqari, uning strukturaviy tashkil etilishi ma'lum turdagi organizmlarga xos bo'lgan bir xil genetik DNK ma'lumotlarini ixtisoslashgan hujayralarda boshqacha ishlatishga imkon beradi.
Elektron mikroskopda xromatinning tasviri boncuklarga o'xshaydi: o'lchami taxminan 10 nm bo'lgan sharsimon qalinlashuvlar, ipga o'xshash ko'priklar bilan ajratilgan. Bunday sharsimon qalinlashuvlar nukleosomalar deb ataladi. Nukleosoma xromatinning strukturaviy birligidir. Har bir nukleosoma har bir nukleosoma yadrosiga 1,75 chap burilish hosil qilish uchun 146-bp supero'ralgan DNK segmentini o'z ichiga oladi. Nukleosoma yadrosi H2A, H2B, H3 va H4 gistonlaridan, har bir turdagi ikkita molekuladan tashkil topgan giston oktameridir (9-rasm), diametri 11 nm va qalinligi 5,7 nm bo'lgan diskka o'xshaydi. Beshinchi giston, H1, nukleosoma yadrosining bir qismi emas va DNKni giston oktameriga o'rash jarayonida ishtirok etmaydi. U qo'sh spiralning nukleosoma yadrosiga kiradigan va undan chiqadigan joylarda DNK bilan aloqa qiladi. Bular interkor (bog'lovchi) DNK bo'limlari bo'lib, ularning uzunligi hujayra turiga qarab 40 dan 50 juft nukleotidgacha o'zgaradi. Natijada, nukleosomalar tarkibiga kiradigan DNK bo'lagining uzunligi ham o'zgarib turadi (186 dan 196 juft nukleotidgacha).
Nukleosomalarda taxminan 90% DNK mavjud, qolganlari bog'lovchilardir. Nukleosomalar "jim" xromatinning bo'laklari va bog'lovchi faol ekanligiga ishoniladi. Biroq, nukleosomalar ochilib, chiziqli bo'lishi mumkin. Ochilmagan nukleosomalar allaqachon faol xromatindir. Bu funktsiyaning tuzilishga bog'liqligini aniq ko'rsatadi. Taxmin qilish mumkinki, globulyar nukleosomalarda xromatin qancha ko'p bo'lsa, u shunchalik faol emas. Shubhasiz, turli hujayralarda tinch xromatinning teng bo'lmagan nisbati bunday nukleosomalar soni bilan bog'liq.
Elektron mikroskopik fotosuratlarda, izolyatsiyalanish shartlariga va cho'zilish darajasiga qarab, xromatin nafaqat qalinlashgan uzun ip - nukleosomalarning "boncuklari" sifatida, balki diametrli qisqaroq va zichroq fibrilla (tola) sifatida ham ko'rinishi mumkin. 30 nm, uning shakllanishi H1 gistonining DNK va H3 gistonining bog'lovchi hududiga bog'langan o'zaro ta'sirida kuzatiladi, bu esa diametri 30 nm bo'lgan solenoid hosil qilish uchun oltita nukleosoma spiralining qo'shimcha burilishiga olib keladi. Bunday holda, giston oqsili bir qator genlarning transkripsiyasiga xalaqit berishi va shu bilan ularning faoliyatini tartibga solishi mumkin.
DNKning yuqorida tavsiflangan gistonlar bilan o'zaro ta'siri natijasida o'rtacha diametri 2 nm va uzunligi 57 nm bo'lgan 186 ta asosiy juftlikdan iborat DNK qo'sh spiralining segmenti 10 nm diametrli spiralga aylanadi. uzunligi 5 nm. Keyinchalik bu spiral diametri 30 nm bo'lgan tolaga siqilganida, kondensatsiya darajasi yana olti baravar ortadi.
Oxir-oqibat, besh gistonli DNK dupleksining qadoqlanishi DNKning 50 marta kondensatsiyasiga olib keladi. Biroq, hatto bunday yuqori darajadagi kondensatsiya ham metafaza xromosomasida DNKning deyarli 50 000 - 100 000 marta siqilishini tushuntira olmaydi. Afsuski, metafaza xromosomasigacha bo'lgan keyingi xromatinli qadoqlash tafsilotlari hali ma'lum emas, shuning uchun biz faqat ushbu jarayonning umumiy xususiyatlarini ko'rib chiqishimiz mumkin.
Xromosomalarda DNKning siqilish darajalari
Har bir DNK molekulasi alohida xromosomaga qadoqlangan. Odamning diploid hujayralarida 46 ta xromosoma mavjud bo'lib, ular hujayra yadrosida joylashgan. Hujayradagi barcha xromosomalar DNKsining umumiy uzunligi 1,74 m ni tashkil qiladi, lekin xromosomalar o'ralgan yadroning diametri millionlab marta kichikdir. Xromosomalar va hujayra yadrosidagi xromosomalardagi DNKning bunday ixcham qadoqlanishi DNK bilan ma'lum bir ketma-ketlikda o'zaro ta'sir qiluvchi turli xil giston va giston bo'lmagan oqsillar tomonidan ta'minlanadi (yuqoriga qarang). Xromosomalardagi DNKni siqish uning chiziqli o'lchamlarini taxminan 10 000 marta - taxminan 5 sm dan 5 mikrongacha kamaytirish imkonini beradi. Siqilishning bir necha darajalari mavjud (10-rasm).


- DNK qo'sh spiral diametri 2 nm va uzunligi bir necha sm bo'lgan manfiy zaryadlangan molekuladir.
- nukleosoma darajasi- xromatin elektron mikroskopda "boncuklar" zanjiri - nukleosomalar - "ip ustida" ko'rinadi. Nukleosoma universal strukturaviy birlik bo'lib, u evromatin va geteroxromatinda, interfaza yadrosi va metafaza xromosomalarida joylashgan.
Siqilishning nukleosoma darajasi maxsus oqsillar - gistonlar tomonidan ta'minlanadi. Sakkizta musbat zaryadlangan giston domenlari manfiy zaryadlangan DNK molekulasi o'ralgan nukleosomaning yadrosini tashkil qiladi. Bu 7 marta qisqarishni beradi, diametri esa 2 dan 11 nm gacha oshadi.
- solenoid darajasi
Xromosomalar tashkil etilishining solenoid darajasi nukleosoma filamentining buralishi va diametri 20-35 nm bo'lgan qalinroq fibrillalar - solenoidlar yoki superbidlarning shakllanishi bilan tavsiflanadi. Solenoid balandligi 11 nm, har bir burilishda taxminan 6-10 nukleosoma mavjud. Solenoidli o'rash superbid o'rashga qaraganda ko'proq deb hisoblanadi, unga ko'ra diametri 20-35 nm bo'lgan xromatin fibrillasi har biri sakkiz nukleosomadan iborat bo'lgan granulalar zanjiri yoki superbidlardir. Solenoid darajasida DNKning chiziqli hajmi 6-10 marta kamayadi, diametri 30 nm gacha oshadi.
- halqa darajasi
Loop darajasi taxminan 30-300 kb gacha bo'lgan halqalarni tashkil etuvchi, ma'lum DNK ketma-ketliklarini taniydigan va bog'laydigan giston joyiga xos bo'lmagan DNKni bog'laydigan oqsillar tomonidan ta'minlanadi. Loop gen ifodasini ta'minlaydi, ya'ni. loop nafaqat strukturaviy, balki funktsional shakllanishdir. Ushbu darajadagi qisqarish 20-30 marta sodir bo'ladi. Diametri 300 nm gacha oshadi. Sitologik preparatlarda amfibiya oositlaridagi "chiroq cho'tkalari" kabi pastadir shaklidagi tuzilmalarni ko'rish mumkin. Ushbu halqalar o'ta o'ralgan ko'rinadi va DNK domenlarini ifodalaydi, ehtimol transkripsiya va xromatin replikatsiyasi birliklariga mos keladi. Maxsus oqsillar halqalarning asoslarini va, ehtimol, ularning ba'zi ichki qismlarini mahkamlaydi. Loopga o'xshash domen tashkiloti metafaza xromosomalarida xromatinning yuqori darajali spiral tuzilmalarga katlanishiga yordam beradi.
- domen darajasi
Xromosomalar tashkil etilishining domen darajasi etarlicha o'rganilmagan. Ushbu darajada halqa domenlarining shakllanishi qayd etilgan - qalinligi 25-30 nm bo'lgan iplar (fibrillalar) tuzilmalari, tarkibida 60% protein, 35% DNK va 5% RNK mavjud bo'lib, hujayra tsiklining barcha bosqichlarida deyarli ko'rinmaydi. mitozdan tashqari va hujayra yadrosi bo'ylab tasodifiy taqsimlanadi. Sitologik preparatlarda amfibiya oositlaridagi "chiroq cho'tkalari" kabi pastadir shaklidagi tuzilmalarni ko'rish mumkin.
Loop domenlari o'z bazasida o'rnatilgan biriktiruvchi saytlar deb ataladigan yadro ichidagi oqsil matritsasiga biriktirilgan bo'lib, ular ko'pincha MAR/SAR ketma-ketliklari deb ataladi (MAR, ingliz matritsasi bilan bog'langan hududdan; SAR, ingliz iskala biriktirma mintaqalaridan) - A/T nukleotid juftlarining yuqori miqdori (>65%) bilan tavsiflangan uzunlikdagi bir necha yuz tayanch juftlikdagi DNK parchalari. Har bir domen replikatsiyaning yagona kelib chiqishiga ega va avtonom o'ta spiral birlik sifatida ishlaydi. Har qanday tsikl domeni ko'plab transkripsiya birliklarini o'z ichiga oladi, ularning ishlashi muvofiqlashtirilgan bo'lishi mumkin - butun domen faol yoki nofaol holatda.
Domen darajasida, ketma-ket xromatinli qadoqlash natijasida DNKning chiziqli o'lchamlari taxminan 200 marta (700 nm) ga kamayadi.
- xromosoma darajasi
Xromosoma darajasida profaza xromosomasining metafaza xromosomasiga kondensatsiyasi giston bo'lmagan oqsillarning eksenel ramkasi atrofida halqa domenlarining siqilishi bilan sodir bo'ladi. Bu superkoillanish hujayradagi barcha H1 molekulalarining fosforlanishi bilan birga kechadi. Natijada, metafaza xromosomasini zich spiralga o'ralgan, zich o'ralgan solenoid halqalar sifatida tasvirlash mumkin. Odatiy odam xromosomasi 2600 tagacha halqadan iborat bo'lishi mumkin. Bunday strukturaning qalinligi 1400 nm (ikki xromatid) ga etadi va DNK molekulasi 104 marta qisqartiriladi, ya'ni. 5 sm cho'zilgan DNK dan 5 mkm gacha.
Xromosomalarning funktsiyalari
Ekstraxromosoma mexanizmlari bilan o'zaro ta'sirda xromosomalar ta'minlaydi
- irsiy ma'lumotlarni saqlash
- uyali aloqani yaratish va saqlash uchun ushbu ma'lumotlardan foydalanish
- irsiy ma'lumotlarni o'qishni tartibga solish
- genetik materialning o'z-o'zidan takrorlanishi
- genetik materialni ona hujayradan qiz hujayralarga o'tkazish.
Xromatin hududi faollashganda, ya'ni. transkripsiya vaqtida undan avval giston H1, keyin esa giston okteti teskari ravishda chiqariladi. Bu xromatin dekondensatsiyasiga, 30 nmli xromatin fibrillasining 10 nmli fibrillaga ketma-ket o'tishiga va keyinchalik erkin DNK bo'limlariga tarqalishiga olib keladi, ya'ni. nukleosoma tuzilishini yo'qotish.
1953 yilda DNK kabi moddaning molekulyar tashkil etilishi printsipi kashf etilgandan so'ng, molekulyar biologiya rivojlana boshladi. Keyinchalik tadqiqot jarayonida olimlar DNK qanday rekombinatsiyalanganligi, uning tarkibi va inson genomimiz qanday tuzilganligini aniqladilar.
Har kuni molekulyar darajada murakkab jarayonlar sodir bo'ladi. DNK molekulasi qanday tuzilgan, u nimadan iborat? Va DNK molekulalari hujayrada qanday rol o'ynaydi? Keling, qo'sh zanjir ichida sodir bo'ladigan barcha jarayonlar haqida batafsil gapiraylik.
Irsiy ma'lumot nima?
Xo'sh, hammasi qaerdan boshlandi? 1868 yilda ular uni bakteriyalar yadrolarida topdilar. 1928 yilda esa N. Koltsov tirik organizm haqidagi barcha genetik ma'lumotlar DNKda shifrlanganligi haqidagi nazariyani ilgari surdi. Keyin J. Uotson va F. Krik 1953 yilda hozirda taniqli DNK spiralining modelini topdilar, buning uchun ular munosib e'tirof va mukofot - Nobel mukofotiga sazovor bo'lishdi.
Qanday bo'lmasin, DNK nima? Ushbu modda 2 ta birlashgan ipdan, aniqrog'i spirallardan iborat. Bunday zanjirning ma'lum ma'lumotlarga ega bo'lgan qismi gen deb ataladi.

DNK qanday oqsillar va qanday tartibda hosil bo'lishi haqidagi barcha ma'lumotlarni saqlaydi. DNK makromolekulasi g'oyat katta hajmdagi ma'lumotlarning moddiy tashuvchisi bo'lib, u alohida g'ishtlar - nukleotidlarning qat'iy ketma-ketligida qayd etiladi. Hammasi bo'lib 4 ta nukleotid mavjud bo'lib, ular kimyoviy va geometrik jihatdan bir-birini to'ldiradi. Fandagi bu to‘ldiruvchi yoki to‘ldiruvchilik tamoyili keyinroq bayon qilinadi. Bu qoida genetik axborotni kodlash va dekodlashda asosiy rol o'ynaydi.
DNK zanjiri nihoyatda uzun bo'lganligi sababli, bu ketma-ketlikda takrorlanishlar mavjud emas. Har bir tirik mavjudot o'ziga xos DNK zanjiriga ega.
DNKning funktsiyalari
Vazifalarga irsiy ma'lumotni saqlash va uni avlodga etkazish kiradi. Ushbu funktsiyasiz turning genomi ming yillar davomida saqlanib qolishi va rivojlanishi mumkin emas edi. Jiddiy gen mutatsiyalariga uchragan organizmlar omon qololmaydi yoki nasl berish qobiliyatini yo'qotadi. Turlarning degeneratsiyasiga qarshi tabiiy himoya shunday sodir bo'ladi.

Yana bir muhim funktsiya saqlangan ma'lumotlarni amalga oshirishdir. Qo'sh zanjirda saqlanadigan ko'rsatmalarsiz hujayra bitta hayotiy oqsilni yarata olmaydi.
Nuklein kislota tarkibi
Nukleotidlarning o'zlari - DNKning qurilish bloklari nimadan iboratligi hozir aniq ma'lum. Ular 3 ta moddani o'z ichiga oladi:
- Ortofosfor kislotasi.
- Azotli asos. Pirimidin asoslari - faqat bitta halqaga ega. Bularga timin va sitozin kiradi. 2 ta halqani o'z ichiga olgan purin asoslari. Bular guanin va adenin.
- saxaroza. DNK tarkibida dezoksiriboza, RNKda riboza mavjud.
Nukleotidlar soni doimo azotli asoslar soniga teng. Maxsus laboratoriyalarda nukleotid parchalanadi va undan azotli asos ajratiladi. Bu nukleotidlarning individual xususiyatlari va ulardagi mumkin bo'lgan mutatsiyalar shunday o'rganiladi.
Irsiy axborotni tashkil etish darajalari
Organizmning 3 darajasi mavjud: genetik, xromosoma va genomik. Yangi oqsil sintezi uchun zarur bo'lgan barcha ma'lumotlar zanjirning kichik qismida - genda joylashgan. Ya'ni, gen axborotni kodlashning eng past va eng oddiy darajasi hisoblanadi.

Genlar, o'z navbatida, xromosomalarga yig'iladi. Irsiy material tashuvchisining bunday tashkil etilishi tufayli xususiyatlar guruhlari ma'lum qonuniyatlarga ko'ra almashadi va bir avloddan ikkinchisiga o'tadi. Shuni ta'kidlash kerakki, tanada aql bovar qilmaydigan miqdordagi genlar mavjud, ammo ma'lumot ko'p marta rekombinatsiya qilinganda ham yo'qolmaydi.
Bir necha turdagi genlar mavjud:
- Funktsional maqsadiga ko'ra 2 turga bo'linadi: strukturaviy va tartibga soluvchi ketma-ketliklar;
- Hujayrada sodir bo'ladigan jarayonlarga ta'siridan kelib chiqib, ular quyidagilardan ajralib turadi: supervital, o'ldiradigan, shartli ravishda o'ldiradigan genlar, shuningdek mutator va antimutator genlar.
Genlar xromosoma bo'ylab chiziqli tartibda joylashgan. Xromosomalarda ma'lumotlar tasodifiy yo'naltirilmaydi, ma'lum bir tartib mavjud. Hatto genlarning joylashuvi yoki joylashishini ko'rsatadigan xarita ham mavjud. Masalan, 18-sonli xromosoma bolaning ko'zining rangi haqidagi ma'lumotlarni shifrlashi ma'lum.
Genom nima? Bu organizm hujayrasidagi nukleotidlar ketma-ketligining butun to'plamiga berilgan nom. Genom individual emas, balki butun turni tavsiflaydi.
Insonning genetik kodi nima?
Gap shundaki, inson rivojlanishining butun ulkan salohiyati allaqachon kontseptsiya davrida yotadi. Zigota rivojlanishi va tug'ilgandan keyin bolaning o'sishi uchun zarur bo'lgan barcha irsiy ma'lumotlar genlarda shifrlangan. DNK bo'limlari irsiy ma'lumotlarning eng asosiy tashuvchilari hisoblanadi.
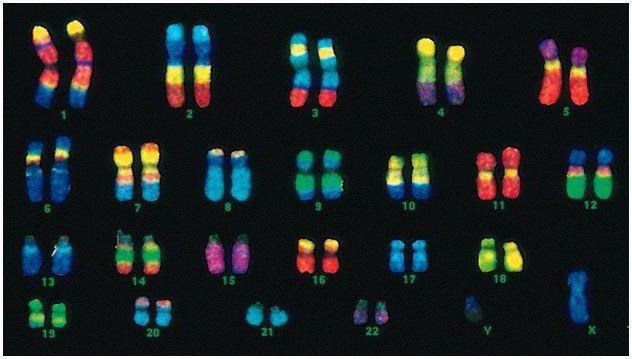
Odamlarda 46 ta xromosoma yoki 22 somatik juft va har bir ota-onadan bitta jinsni aniqlovchi xromosoma mavjud. Ushbu diploid xromosomalar to'plami insonning butun jismoniy ko'rinishini, uning aqliy va jismoniy qobiliyatlarini va kasalliklarga moyilligini kodlaydi. Somatik xromosomalar tashqi tomondan farqlanmaydi, ammo ular turli xil ma'lumotlarni o'z ichiga oladi, chunki ulardan biri otadan, ikkinchisi onadan.
Erkak kodi ayol kodidan xromosomalarning oxirgi juftligi - XYda farqlanadi. Ayol diploid to'plami oxirgi juftlik, XX. Erkaklar biologik onasidan bitta X xromosomasini oladi, keyin u qizlariga o'tadi. Jinsiy Y xromosomasi o'g'illarga o'tadi.
Inson xromosomalari hajmi jihatidan juda farq qiladi. Masalan, xromosomalarning eng kichik juftligi No17. Va eng katta juftlik 1 va 3.
Odamlarda qo'sh spiralning diametri atigi 2 nm. DNK shu qadar mahkam o'ralganki, u hujayraning kichik yadrosiga sig'adi, garchi burilmagan bo'lsa, uning uzunligi 2 metrgacha bo'ladi. Spiral uzunligi yuz millionlab nukleotidlarni tashkil qiladi.
Genetik kod qanday uzatiladi?
Xo'sh, DNK molekulalari hujayra bo'linishida qanday rol o'ynaydi? Genlar - irsiy ma'lumotlarning tashuvchilari - tananing har bir hujayrasida joylashgan. Ko'p jonzotlar o'z kodlarini qiz organizmiga o'tkazish uchun o'zlarining DNKlarini ikkita bir xil spiralga bo'lishadi. Bu replikatsiya deb ataladi. Replikatsiya jarayonida DNK ochiladi va maxsus "mashinalar" har bir ipni to'ldiradi. Genetik spiral ikkiga bo'lingandan so'ng, yadro va barcha organellalar, so'ngra butun hujayra bo'linishni boshlaydi.
Ammo odamlarda genlarni uzatishning boshqa jarayoni bor - jinsiy. Ota va onaning xususiyatlari aralash, yangi genetik kod ikkala ota-onadan ma'lumotni o'z ichiga oladi.
Irsiy axborotni saqlash va uzatish DNK spiralining murakkab tashkil etilishi tufayli mumkin. Axir, aytganimizdek, oqsillarning tuzilishi genlarda shifrlangan. Kontseptsiya vaqtida yaratilganidan so'ng, bu kod hayot davomida o'zini ko'rsatadi. Karyotip (xromosomalarning shaxsiy to'plami) organ hujayralarining yangilanishi paytida o'zgarmaydi. Ma'lumotni uzatish jinsiy gametalar - erkak va ayol yordamida amalga oshiriladi.
Faqat bitta RNK zanjirini o'z ichiga olgan viruslar o'z ma'lumotlarini avlodlariga etkazishga qodir emas. Shuning uchun ular ko'payish uchun odam yoki hayvon hujayralariga muhtoj.
Irsiy axborotni amalga oshirish
Hujayra yadrosida doimo muhim jarayonlar sodir bo'ladi. Xromosomalarda qayd etilgan barcha ma'lumotlar aminokislotalardan oqsillarni yaratish uchun ishlatiladi. Ammo DNK zanjiri hech qachon yadrodan chiqmaydi, shuning uchun u boshqa muhim birikma - RNK yordamiga muhtoj. Aynan RNK yadro membranasiga kirib, DNK zanjiri bilan ta'sir o'tkazishga qodir.
DNK va 3 turdagi RNKning o'zaro ta'siri orqali barcha kodlangan ma'lumotlar amalga oshiriladi. Irsiy axborotni amalga oshirish qaysi darajada sodir bo'ladi? Barcha o'zaro ta'sirlar nukleotidlar darajasida sodir bo'ladi. Messenger RNK DNK zanjirining bir qismini nusxalaydi va bu nusxani ribosomaga olib keladi. Bu erda nukleotidlardan yangi molekulaning sintezi boshlanadi.
mRNK zanjirning kerakli qismini nusxalashi uchun spiral ochiladi va keyin qayta kodlash jarayoni tugagandan so'ng yana tiklanadi. Bundan tashqari, bu jarayon bir vaqtning o'zida 1 xromosomaning 2 tomonida sodir bo'lishi mumkin.
To'ldiruvchilik printsipi
Ular 4 ta nukleotiddan iborat - adenin (A), guanin (G), sitozin (C), timin (T). Ular komplementarlik qoidasiga ko'ra vodorod bog'lari bilan bog'langan. E. Chargaffning ishi bu qoidani o'rnatishga yordam berdi, chunki olim bu moddalarning xatti-harakatlarida ba'zi naqshlarni payqadi. E. Chargaff adenin va timinning molyar nisbati birga teng ekanligini aniqladi. Va xuddi shu tarzda, guanin va sitozin nisbati har doim birga teng.
Uning ishiga asoslanib, genetiklar nukleotidlarning o'zaro ta'siri uchun qoidani shakllantirdilar. Komplementarlik qoidasi shuni ko'rsatadiki, adenin faqat timin bilan, guanin esa faqat sitozin bilan birlashadi. Spiralning dekodlanishi va ribosomada yangi oqsil sintezi jarayonida bu almashinish qoidasi transfer RNKga biriktirilgan kerakli aminokislotalarni tezda topishga yordam beradi.
RNK va uning turlari
Irsiy ma'lumot nima? DNKning qo'sh zanjiridagi nukleotidlar. RNK nima? Uning ishi nima? RNK yoki ribonuklein kislotasi DNKdan ma'lumot olish, uni dekodlash va komplementarlik printsipiga asoslanib, hujayralar uchun zarur bo'lgan oqsillarni yaratishga yordam beradi.
Hammasi bo'lib 3 xil RNK mavjud. Ularning har biri o'z vazifasini qat'iy bajaradi.
- Axborot (mRNK), yoki matritsa deb ham ataladi. U to'g'ridan-to'g'ri hujayraning markaziga, yadroga kiradi. Xromosomalardan birida oqsil hosil qilish uchun zarur bo'lgan genetik materialni topadi va qo'sh ipning bir tomonini ko'chiradi. Nusxa ko'chirish bir-birini to'ldirish tamoyiliga ko'ra yana sodir bo'ladi.
- Transport kichik molekula bo'lib, uning bir tomonida nukleotid dekoderlari, ikkinchi tomonida esa asosiy kodga mos keladigan aminokislotalar mavjud. tRNKning vazifasi uni "ustaxonaga", ya'ni kerakli aminokislotalarni sintez qiladigan ribosomaga etkazishdir.
- rRNK ribosomadir. U ishlab chiqarilgan protein miqdorini nazorat qiladi. U 2 qismdan iborat - aminokislota va peptid bo'limi.
Dekodlashning yagona farqi shundaki, RNKda timin yo'q. Bu erda timin o'rniga urasil mavjud. Ammo keyin, oqsil sintezi jarayonida, tRNK hali ham barcha aminokislotalarni to'g'ri o'rnatadi. Agar ma'lumotni dekodlashda biron bir nosozlik yuzaga kelsa, u holda mutatsiya sodir bo'ladi.
Buzilgan DNK molekulasini tiklash
Shikastlangan er-xotin ipni tiklash jarayoni ta'mirlash deb ataladi. Ta'mirlash jarayonida shikastlangan genlar olib tashlanadi.

Keyin elementlarning kerakli ketma-ketligi aniq takrorlanadi va zanjirning olib tashlangan joyidan bir xil joyga kesiladi. Bularning barchasi maxsus kimyoviy moddalar - fermentlar tufayli sodir bo'ladi.
Nima uchun mutatsiyalar paydo bo'ladi?
Nima uchun ba'zi genlar mutatsiyaga kirishadi va o'z vazifalarini bajarishni to'xtatadi - hayotiy irsiy ma'lumotlarni saqlash? Bu dekodlashda xatolik tufayli yuzaga keladi. Masalan, adenin tasodifan timin bilan almashtirilsa.
Xromosoma va genomik mutatsiyalar ham mavjud. Xromosoma mutatsiyalari irsiy ma'lumotlarning bo'limlari yo'qolganda, ko'payganda yoki hatto boshqa xromosomaga ko'chirilganda sodir bo'ladi.

Genomik mutatsiyalar eng jiddiy hisoblanadi. Ularning sababi xromosomalar sonining o'zgarishi. Ya'ni, karyotipda juft - diploid to'plam o'rniga triploid to'plam mavjud bo'lganda.
Triploid mutatsiyaning eng mashhur misoli Daun sindromi bo'lib, unda xromosomalarning shaxsiy to'plami 47. Bunday bolalarda 21-juft o'rnida 3 ta xromosoma hosil bo'ladi.
Poliploidiya deb ataladigan ma'lum mutatsiya ham mavjud. Ammo poliploidiya faqat o'simliklarda uchraydi.
DNK molekulasi qo'sh spiral hosil qiluvchi ikkita zanjirdan iborat. Uning tuzilishi birinchi marta 1953 yilda Frensis Krik va Jeyms Uotson tomonidan shifrlangan.
Dastlab, bir-birining atrofida o'ralgan bir juft nukleotid zanjiridan iborat DNK molekulasi nima uchun bunday o'ziga xos shaklga ega ekanligi haqida savollar tug'dirdi. Olimlar bu hodisani komplementarlik deb atashadi, ya'ni uning iplarida faqat ma'lum nukleotidlar bir-biriga qarama-qarshi bo'lishi mumkin. Masalan, adenin doimo timinga, guanin esa har doim sitozinga qarama-qarshidir. DNK molekulasining bu nukleotidlari komplementar deb ataladi.
Sxematik ravishda u quyidagicha tasvirlangan:
T - A
C - G
Bu juftliklar aminokislotalarning joylashish tartibini belgilaydigan kimyoviy nukleotid bog'lanish hosil qiladi. Birinchi holda, u biroz zaifroq. C va G o'rtasidagi aloqa kuchliroq. Komplementar bo'lmagan nukleotidlar bir-biri bilan juftlik hosil qilmaydi.
Bino haqida
Demak, DNK molekulasining tuzilishi alohida. Uning bunday shakli bor: haqiqat shundaki, nukleotidlar soni juda ko'p va uzun zanjirlarni joylashtirish uchun juda ko'p joy kerak. Aynan shuning uchun zanjirlar spiral burilish bilan tavsiflanadi. Ushbu hodisa spiralizatsiya deb ataladi, bu iplarni taxminan besh-olti marta qisqartirish imkonini beradi.
Tana bunday turdagi ba'zi molekulalarni juda faol ishlatadi, boshqalari kamdan-kam hollarda. Ikkinchisi, spiralizatsiyadan tashqari, superspiralizatsiya kabi "ixcham qadoqlash" dan ham o'tadi. Va keyin DNK molekulasining uzunligi 25-30 marta qisqaradi.

Molekulaning "qadoqlanishi" nima?
Supercoiling jarayoni giston oqsillarini o'z ichiga oladi. Ular ipning g'altak yoki novda tuzilishi va ko'rinishiga ega. Ularga spirallangan iplar o'raladi, ular darhol "ixcham qadoqlangan" va kam joy egallaydi. U yoki bu ipni ishlatish zarurati tug'ilganda, u g'altakdan, masalan, giston oqsilidan ochiladi va spiral ikkita parallel zanjirga aylanadi. DNK molekulasi bu holatda bo'lganda, undan kerakli genetik ma'lumotlarni o'qish mumkin. Biroq, bitta shart mavjud. Agar DNK molekulasining tuzilishi burilmagan shaklga ega bo'lsa, ma'lumot olish mumkin. O'qish mumkin bo'lgan xromosomalar evromatinlar deb ataladi va agar ular o'ta o'ralgan bo'lsa, ular allaqachon geteroxromatinlardir.
Nuklein kislotalar
Nuklein kislotalar, oqsillar kabi, biopolimerlardir. Asosiy funktsiya irsiy (genetik ma'lumotni) saqlash, amalga oshirish va uzatishdir. Ular ikki xil bo'ladi: DNK va RNK (deoksiribonuklein va ribonuklein). Ulardagi monomerlar nukleotidlar bo'lib, ularning har birida fosfor kislotasi qoldig'i, besh uglerodli shakar (dezoksiriboza / riboza) va azotli asos mavjud. DNK kodi 4 turdagi nukleotidlarni o'z ichiga oladi - adenin (A) / guanin (G) / sitozin (C) / timin (T). Ular tarkibidagi azotli asosda farqlanadi.
DNK molekulasida nukleotidlar soni juda katta bo'lishi mumkin - bir necha mingdan o'nlab va yuzlab millionlargacha. Bunday ulkan molekulalarni elektron mikroskop orqali tekshirish mumkin. Bunday holda, siz bir-biriga nukleotidlarning azotli asoslarining vodorod aloqalari bilan bog'langan polinukleotid zanjirlarining qo'sh zanjirini ko'rishingiz mumkin.

Tadqiqot
Tadqiqotlar davomida olimlar DNK molekulalarining turlari turli tirik organizmlarda farqlanishini aniqladilar. Shuningdek, bitta zanjirning guanini faqat sitozin bilan, timin esa adenin bilan bog'lanishi mumkinligi aniqlandi. Bir zanjirdagi nukleotidlarning joylashishi parallel zanjirga to'g'ri keladi. Polinukleotidlarning bu to'ldiruvchiligi tufayli DNK molekulasi ikki baravar ko'payib, o'z-o'zini ko'paytirishga qodir. Ammo birinchi navbatda, qo'shilgan nukleotidlarni yo'q qiladigan maxsus fermentlar ta'siri ostida qo'shimcha zanjirlar ajralib chiqadi va keyin ularning har birida etishmayotgan zanjirning sintezi boshlanadi. Bu har bir hujayrada ko'p miqdorda bo'lgan erkin nukleotidlar tufayli yuzaga keladi. Natijada, "ona molekulasi" o'rniga tarkibi va tuzilishi jihatidan bir xil bo'lgan ikkita "qizi" hosil bo'ladi va DNK kodi asl nusxaga aylanadi. Bu jarayon hujayra bo'linishining kashshofidir. U barcha irsiy ma'lumotlarning ona hujayralaridan qiz hujayralariga, shuningdek, barcha keyingi avlodlarga uzatilishini ta'minlaydi.
Gen kodi qanday o'qiladi?
Bugungi kunda nafaqat DNK molekulasining massasi hisoblab chiqiladi, balki ilgari olimlar uchun mavjud bo'lmagan murakkabroq ma'lumotlarni ham topish mumkin. Masalan, organizm o'z hujayrasidan qanday foydalanishi haqidagi ma'lumotlarni o'qishingiz mumkin. Albatta, dastlab bu ma'lumot kodlangan shaklda bo'lib, ma'lum bir matritsa shakliga ega va shuning uchun uni RNK bo'lgan maxsus tashuvchiga o'tkazish kerak. Ribonuklein kislota yadro membranasi orqali hujayra ichiga kirib, ichidagi kodlangan ma'lumotlarni o'qiy oladi. Shunday qilib, RNK yadrodan hujayraga yashirin ma'lumotlarning tashuvchisi bo'lib, uning DNKdan farqi shundaki, unda dezoksiriboza o'rniga riboza, timin o'rniga urasil mavjud. Bundan tashqari, RNK bir zanjirli.

RNK sintezi
DNKni chuqur tahlil qilish shuni ko'rsatdiki, RNK yadrodan chiqqandan so'ng u sitoplazmaga kiradi va u erda matritsa sifatida ribosomalarga (maxsus ferment tizimlari) birlashishi mumkin. Qabul qilingan ma'lumotlarga asoslanib, ular protein aminokislotalarining tegishli ketma-ketligini sintez qilishlari mumkin. Ribosoma triplet kodidan hosil qiluvchi oqsil zanjiriga qaysi turdagi organik birikmalar biriktirilishi kerakligini bilib oladi. Har bir aminokislota uni kodlaydigan o'ziga xos tripletga ega.
Zanjirning shakllanishi tugagandan so'ng, u o'ziga xos fazoviy shaklga ega bo'ladi va o'zining gormonal, qurilish, fermentativ va boshqa funktsiyalarini bajarishga qodir bo'lgan oqsilga aylanadi. Har qanday organizm uchun bu gen mahsulotidir. Undan genlarning barcha turdagi sifatlari, xossalari va ko'rinishlari aniqlanadi.

Genlar
Sekvensiya jarayonlari birinchi navbatda DNK molekulasi tarkibida qancha gen borligi haqida ma'lumot olish uchun ishlab chiqilgan. Va tadqiqot olimlarga bu masalada katta yutuqlarga erishishga imkon bergan bo'lsa-da, ularning aniq sonini bilish hali mumkin emas.
Bir necha yil oldin, DNK molekulalarida taxminan 100 ming gen borligi taxmin qilingan edi. Biroz vaqt o'tgach, bu raqam 80 mingga kamaydi va 1998 yilda genetiklar bitta DNKda atigi 50 ming gen mavjudligini ta'kidladilar, bu umumiy DNK uzunligining atigi 3 foizini tashkil qiladi. Ammo genetiklarning so'nggi xulosalari hayratlanarli edi. Endi ular genomda bu birliklarning 25-40 mingini o'z ichiga oladi, deb da'vo qilmoqdalar. Ma'lum bo'lishicha, xromosoma DNKsining atigi 1,5 foizi oqsillarni kodlash uchun javobgardir.
Tadqiqotlar shu bilan to‘xtab qolmadi. Parallel genetik muhandislik mutaxassislari guruhi bitta molekuladagi genlar soni roppa-rosa 32 ming ekanligini aniqladi. Ko'rib turganingizdek, aniq javob olish hali ham mumkin emas. Qarama-qarshiliklar juda ko'p. Barcha tadqiqotchilar faqat o'z natijalariga tayanadilar.
Evolyutsiya bo'lganmi?
Molekulaning evolyutsiyasi haqida hech qanday dalil yo'qligiga qaramay (chunki DNK molekulasining tuzilishi mo'rt va kichik o'lchamli), olimlar hali ham bitta taxminni ilgari surdilar. Laboratoriya ma'lumotlariga asoslanib, ular quyidagi versiyani aytishdi: paydo bo'lishining dastlabki bosqichida molekula qadimgi okeanlarda topilgan 32 tagacha aminokislotalarni o'z ichiga olgan oddiy o'z-o'zini ko'paytiruvchi peptid shakliga ega edi.
O'z-o'zini ko'paytirishdan so'ng, tabiiy tanlanish kuchlari tufayli molekulalar o'zlarini tashqi elementlardan himoya qilish qobiliyatiga ega bo'ldilar. Ular uzoqroq yashay boshladilar va ko'proq miqdorda ko'paya boshladilar. O'zlarini lipid pufagida topib olgan molekulalar o'zlarini ko'paytirish uchun barcha imkoniyatlarga ega edi. Bir qator ketma-ket tsikllar natijasida lipid pufakchalari hujayra membranalari, keyin esa taniqli zarrachalar shakliga ega bo'ldi. Shuni ta'kidlash kerakki, bugungi kunda DNK molekulasining har qanday bo'limi murakkab va aniq ishlaydigan tuzilma bo'lib, uning barcha xususiyatlarini olimlar hali to'liq o'rganmagan.

Zamonaviy dunyo
Yaqinda isroillik olimlar soniyada trillionlab operatsiyalarni bajara oladigan kompyuterni yaratdilar. Bugungi kunda bu Yerdagi eng tez avtomobil. Butun sir shundaki, innovatsion qurilma DNK tomonidan quvvatlanadi. Professorlarning aytishicha, yaqin kelajakda bunday kompyuterlar hatto energiya ishlab chiqarishga qodir.
Bir yil avval Rehovotdagi (Isroil) Veysman instituti mutaxassislari molekulalar va fermentlardan iborat dasturlashtiriladigan molekulyar hisoblash mashinasi yaratilishini e’lon qilishdi. Ular silikon mikrochiplarni almashtirdilar. Bugungi kunga kelib, jamoa yanada oldinga siljidi. Endi faqat bitta DNK molekulasi kompyuterni kerakli ma'lumotlar va kerakli yoqilg'i bilan ta'minlay oladi.
Biokimyoviy "nanokompyuterlar" fantastika emas, ular tabiatda allaqachon mavjud va har bir tirik mavjudotda namoyon bo'ladi. Ammo ko'pincha ular odamlar tomonidan boshqarilmaydi. Aytaylik, "Pi" sonini hisoblash uchun odam hali biron bir o'simlikning genomida operatsiya qila olmaydi.
Ma'lumotlarni saqlash/qayta ishlash uchun DNKdan foydalanish g'oyasi birinchi marta olimlarning ongiga 1994 yilda kelgan. Aynan o'sha paytda molekula oddiy matematik masalani hal qilish uchun ishlatilgan. O'shandan beri bir qator tadqiqot guruhlari DNK kompyuterlari bilan bog'liq turli loyihalarni taklif qilishdi. Ammo bu erda barcha urinishlar faqat energiya molekulasiga asoslangan edi. Bunday kompyuterni oddiy ko'z bilan ko'ra olmaysiz, u probirkadagi suvning shaffof eritmasiga o'xshaydi. Unda mexanik qismlar yo'q, faqat trillionlab biomolekulyar qurilmalar - va bu faqat bir tomchi suyuqlikda!
Inson DNKsi
Odamlar 1953-yilda, olimlar birinchi marta ikki zanjirli DNK modelini dunyoga ko'rsatishga muvaffaq bo'lganida, inson DNKsining turi haqida bilishgan. Buning uchun Kirk va Uotson Nobel mukofotiga sazovor bo'lishdi, chunki bu kashfiyot 20-asrda asos bo'ldi.
Vaqt o'tishi bilan, albatta, ular tuzilgan inson molekulasi nafaqat taklif qilingan versiyadagi kabi ko'rinishi mumkinligini isbotladilar. Batafsilroq DNK tahlilini o'tkazgandan so'ng, ular A-, B- va chap qo'l Z- shaklini topdilar. A shakli ko'pincha istisno hisoblanadi, chunki u namlik etishmasligi bilan hosil bo'ladi. Ammo bu faqat laboratoriya tadqiqotlarida mumkin; tabiiy muhit uchun bu anomaldir, tirik hujayrada bunday jarayon bo'lishi mumkin emas.
B-shakli klassik bo'lib, ikki tomonlama o'ng qo'l zanjiri sifatida tanilgan, ammo Z-shakli nafaqat chapga qarama-qarshi yo'nalishda buralibgina qolmay, balki ko'proq zigzag ko'rinishga ega. Olimlar G-quadrupleks shaklini ham aniqladilar. Uning tuzilishi 2 emas, balki 4 ta ipdan iborat. Genetiklarning fikriga ko'ra, bu shakl guaninning ortiqcha miqdori bo'lgan joylarda paydo bo'ladi.

Sun'iy DNK
Bugungi kunda sun'iy DNK allaqachon mavjud bo'lib, u haqiqiyning bir xil nusxasi; u tabiiy qo'sh spiralning tuzilishiga mukammal tarzda amal qiladi. Ammo, asl polinukleotiddan farqli o'laroq, sun'iyda faqat ikkita qo'shimcha nukleotid mavjud.
Dublyaj haqiqiy DNKning turli xil tadqiqotlari natijasida olingan ma'lumotlar asosida yaratilganligi sababli, uni nusxalash, o'z-o'zini ko'paytirish va rivojlantirish ham mumkin. Mutaxassislar taxminan 20 yil davomida shunday sun'iy molekulani yaratish ustida ishlamoqda. Natijada genetik kodni tabiiy DNK kabi ishlata oladigan ajoyib ixtiro.
Mavjud to'rtta azotli asosga genetiklar tabiiy asoslarning kimyoviy modifikatsiyasi natijasida yaratilgan ikkita qo'shimcha asosni qo'shdilar. Tabiiy DNKdan farqli o'laroq, sun'iy DNK juda qisqa bo'lib chiqdi. U faqat 81 ta asosiy juftlikni o'z ichiga oladi. Biroq, u ham ko'payadi va rivojlanadi.
Sun'iy ravishda olingan molekulaning replikatsiyasi polimeraza zanjiri reaktsiyasi tufayli sodir bo'ladi, ammo hozircha bu mustaqil ravishda emas, balki olimlarning aralashuvi bilan sodir bo'ladi. Ular mustaqil ravishda aytilgan DNKga kerakli fermentlarni qo'shib, uni maxsus tayyorlangan suyuq muhitga joylashtiradilar.
Yakuniy natija
DNK rivojlanishining jarayoni va yakuniy natijasiga mutatsiyalar kabi turli omillar ta'sir ko'rsatishi mumkin. Bu tahlil natijasi ishonchli va ishonchli bo'lishi uchun materiya namunalarini o'rganish zarurligini keltirib chiqaradi. Bunga misol qilib, otalik testini keltirish mumkin. Ammo mutatsiya kabi hodisalar kamdan-kam uchraydiganidan quvonmay ilojimiz yo'q. Shunga qaramay, tahlil asosida aniqroq ma'lumot olish uchun materiya namunalari doimo qayta tekshiriladi.

O'simlik DNKsi
Yuqori ketma-ketlik texnologiyalari (HTS) tufayli genomika sohasida inqilob amalga oshirildi - o'simliklardan DNK olish ham mumkin. Albatta, o‘simlik materialidan yuqori sifatli molekulyar og‘irlikdagi DNKni olish mitoxondriya va xloroplast DNK nusxalarining ko‘pligi, shuningdek, polisaxaridlar va fenolik birikmalarning yuqori darajasi tufayli ma’lum qiyinchiliklar tug‘diradi. Bu holda biz ko'rib chiqayotgan strukturani izolyatsiya qilish uchun turli usullar qo'llaniladi.
DNKdagi vodorod aloqasi
DNK molekulasidagi vodorod aloqasi elektronegativ atomga biriktirilgan musbat zaryadlangan vodorod atomi o'rtasida hosil bo'lgan elektromagnit tortishish uchun javobgardir. Bu dipol o'zaro ta'sir kimyoviy bog'lanish mezoniga javob bermaydi. Ammo u molekulalararo yoki molekulaning turli qismlarida, ya'ni intramolekulyar ravishda sodir bo'lishi mumkin.

Vodorod atomi bog'ning donori bo'lgan elektronegativ atomga biriktiriladi. Elektromanfiy atom azot, ftor yoki kislorod bo'lishi mumkin. U - markazsizlashtirish orqali - vodorod yadrosidan elektron bulutini o'ziga tortadi va vodorod atomini (qisman) musbat zaryad qiladi. H ning kattaligi boshqa molekulalar va atomlarga nisbatan kichik bo'lgani uchun zaryad ham kichikdir.
DNK dekodlash
DNK molekulasini dekodlashdan oldin olimlar birinchi navbatda juda ko'p hujayralarni olishadi. Eng aniq va muvaffaqiyatli ish uchun ulardan bir millionga yaqini kerak bo'ladi. Tadqiqot davomida olingan natijalar doimiy ravishda taqqoslanadi va qayd etiladi. Bugungi kunda genomni dekodlash endi kamdan-kam uchraydigan narsa emas, balki foydalanish mumkin bo'lgan protseduradir.

Albatta, bitta hujayraning genomini ochish amaliy bo'lmagan mashqdir. Bunday tadqiqotlar davomida olingan ma'lumotlar olimlarni qiziqtirmaydi. Ammo shuni tushunish kerakki, hozirda mavjud bo'lgan barcha dekodlash usullari, ularning murakkabligiga qaramay, etarlicha samarali emas. Ular faqat DNKning 40-70 foizini o'qishga imkon beradi.
Biroq, Garvard professorlari yaqinda genomning 90% ni shifrlash mumkin bo'lgan usulni e'lon qilishdi. Texnika ajratilgan hujayralarga primer molekulalarni qo'shishga asoslangan bo'lib, ular yordamida DNK replikatsiyasi boshlanadi. Ammo bu usulni ham muvaffaqiyatli deb bo'lmaydi, uni fanda ochiq qo'llashdan oldin uni hali ham takomillashtirish kerak.
